МЕТОДЫ ИЗУЧЕНИЯ
Обычно выделяют следующие этапы эволюции человека:
1. Древнейшие стадии гоминизации - происхождение рода Homo.
2. Эволюция рода Homo до возникновения современного человека.
3. Эволюция современного человека.
Первый этап антропогенеза есть чисто биологическая эволюция. На втором этапе к элементарным факторам биологической эволюции подключается действие социального фактора, который на третьем этапе является доминирующим. Методологические подходы к изучению разных этапов антропогенеза различны.
Изучение 1-го этапа производят методами палеонтологии и сравнительной анатомии. В связи с появлением элементов материальной культуры 2-й этап изучают также методами археологии. На 3-м этапе эволюционные события происходят в основном на молекулярно-генетическом уровне и проявляются на популяционном уровне. Основными способами изучения эволюции современного человека поэтому являются биохимический, цитогенетический и популяционно-статистический методы.
Изучение первых двух этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры. Для определения абсолютного возраста ископаемых остатков человека и его предков широко используют физические методы, в частности радиометрические. С помощью масс-спектрометров определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца. Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод абсолютной датировки ископаемых остатков: чем меньше коллагена содержится в образце, тем более велика его древность.
В последнее время в антропологии все более активно применяют методы иммунологии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее. Для определения прямого родства организмов друг с другом используют иммунологический метод, основанный на изучении иммунологических реакций антиген - антитело. Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов. Из современных человекообразных обезьян к человеку иммунологически наиболее близок шимпанзе, наиболее далеко от человека отстоит орангутан.
Иммунологическим методом было обнаружено, что белки рамапитека, человекообразной обезьяны Южной Азии (абс. возраст 13 млн. лет), более сходны с белками орангутана, чем человека и шимпанзе. Эти данные вместе с результатами морфологических и палеонтологических сопоставлений заставили отказаться от представления о том, что рамапитек является прямым предком человека, и связать его с эволюционной линией орангутана. Из этого следует, что разделение человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн. лет назад.
В последние годы антропогенез эффективно изучают также биомолекулярными методами. В основе этих методов изучения эволюции лежит допущение, что мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут более"сходными друг с другом, чем имевшие его очень давно. Сущность биомолекулярных методов состоит в использовании данных, полученных при сопоставлении белков и нуклеиновых кислот организмов разных видов для определения их родства и древности соответствующих филогенетических ветвей. При этом считают, что степень различий в аминокислотном составе белков и в нуклеотидных последовательностях ДНК позволяет судить о времени расхождения сравниваемых видов от предковой формы. Естественно, что оценивать степень родства и давность расхождения филогенетических ветвей можно лишь по накоплению нейтральных мутаций в геноме, так как вредные мутации быстро элиминируются из генофондов. В действительности нельзя считать большинство мутаций селективно нейтральными.
Расчеты показывают, что при нейтральности мутаций скорость их накопления постоянна только при измерении времени в количестве поколений, а не в годах. Так как продолжительность жизни поколений у различных видов различна, то и скорости накопления различий нуклеотидных последовательностей будут более велики у короткоживущих видов по сравнению с долгоживущими. Кроме того, фактическая скорость эволюции может значительно варьировать в различные временные интервалы в различных группах и по разным признакам. В определении применимости методов молекулярной биологии имеет значение возможность возникновения конвергентного сходства молекул, причем вероятность его повышается с увеличением изучаемых временных интервалов. Эволюция генов и белков часто может опережать реальное расхождение популяций, в первую очередь за счет адаптивного генетического полиморфизма. Однако молекулярно-биологические методы применимы для оценки родства и времени дивергенции в качестве приблизительных «молекулярных часов» при сравнении средних скоростей замен нуклеотидов в ДНК в целом и аминокислот во многих белках за длительные интервалы времени.
Биомолекулярный подход - лишь один из путей определения эволюционных расстояний, который работает только наряду с классическими методами палеонтологии и антропологии, причем в результатах при этом возможны серьезные расхождения. Так, при изучении скелета человека, обнаруженного на территории Эквадора, по данным радиоуглеродного и аминокислотного анализа была установлена его древность в 28 тыс. лет. При использовании же коллагенового анализа возраст того же скелета оказался датированным не более чем в 2,5 тыс. лет.
В антропологии применяют несколько методов оценки степени дифференцированности таксонов, дополняющих друг друга: гибридизация ДНК, определение аминокислотных последовательностей белков, изучение генов гистосовместимости и т.д. Наиболее достоверные данные получены путем гибридизации ДНК, так как скорость эволюции ДНК в целом более постоянна, чем скорость изменения белков. Гибридизация ДНК показала, что момент дивергенции эволюционных ветвей человека и шимпанзе наступил 6,5-6,7 млн. лет назад (рис. 15.2).
Однако между эволюцией структуры генома в виде накопления генных мутаций и морфофизиологической эволюцией часто нет прямой зависимости (рис. 15.3). Это может быть связано с тем, что в формировании практически всех сложных фенотипических признаков принимают участие различные генные системы. Таким образом, скорость эволюции белков у двух разных родственных видов может быть одинакова, а скорость эволюции в целом, оцененная по комплексу фенотипических признаков, при этом оказывается различной. Сравнение аминокислотных последовательностей белков шимпанзе и человека привело к выводу, что около 99% их белков абсолютно идентичны. Из этого следует, что и структурные гены человека и шимпанзе сходны в наибольшей степени. С чем же связаны столь значительные морфофизиологические отличия обоих видов? Можно предполагать, что это зависит от различного распределения белков в клетках организма в процессе развития, что, в свою очередь, определяется различиями программы считывания сходной наследственной информации во времени и пространстве. Иными словами, это может быть обусловлено эволюционными преобразованиями в большей степени не структурных, а регуляторных генов.
Рис. 15.2. Среднее время дивергенции высших приматов по часам ДНК
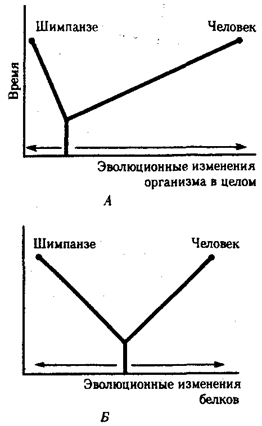
Рис. 15.3. Соотношение скорости морфофизиологической (А ) и молекулярной (Б ) эволюции
Поскольку изучение хромосомного материала возможно только у ныне живущих организмов, применение цитогенетического метода ограничивается современным человеком и человекообразными обезьянами. Дифференциальная окраска хромосом позволяет не только сопоставлять хромосомы разных видов приматов и человека и изучать хромосомный полиморфизм современного человека, но и решать некоторые вопросы эволюции.
Выяснено, что кариотип человекообразных обезьян отличается по количеству хромосом от кариотипа человека на одну пару (23 пары хромосом человека и 24 пары шимпанзе). У человека и шимпанзе идентичны 13 пар хромосом. Хромосома 2-й пары человека точно соответствует двум соединенным хромосомам шимпанзе, а остальные хромосомы отличаются друг от друга незначительно. Так, хромосома 5-й пары шимпанзе соответствует такой же хромосоме человека, но небольшой ее перицентрический участок инвертирован на 180° по сравнению с человеческой хромосомой. Инверсии такого рода обнаружены в кариотипах человека и шимпанзе еще в восьми хромосомах. Эти данные вместе с указаниями на сходство белков человека и шимпанзе свидетельствуют об их значительной эволюционной близости.
Сопоставление кариотипов людей, происходящих из разных популяций, приводит к выводу о полиморфизме хромосом, в первую очередь по размерам гетерохроматиновых участков. Наследуемость индивидуальных вариаций хромосом и их неравномерное распределение в разных популяциях (в частности, расовые различия по размерам длинного плеча Y-хромосомы) делают возможным популяционно-цитогенетический подход в изучении эволюции современного человека.
Эволюционистские круги были потрясены результатами последней передатировки ископаемых останков Homo erectus ("человека прямоходящего"), найденных на острове Ява в Нгандонгском пласте у реки Соло. Считалось, что этот предполагаемый предок современного человека вымер очень давно. Новая датировка ископаемых - максимальный возраст 46 тыс. лет и наиболее вероятный возраст 27 тыс. лет - говорит о том, что Homo Erectus не исчез с появлением анатомически современного человека (Homo sapiens
), а еще долго сосуществовал с ним. Эти данные идут вразрез с существующей концепцией эволюции человека.
Результаты исследований, проведенных группой ученых во главе с Карлом Свишером III и Г.Х.Кертисом из Геохронологического центра Беркли, были опубликованы в журнале "Science" за 13 декабря 1996 г. Была произведена датировка двух отложений ископаемых на Яве - Соло (Нгандонг) и Самбунгмакан - с помощью двух методов датирования, электрон-спинового резонанса и масс-спектрометрией серий U. Проект Свишера и его группы был направлен на поиск новых данных об одной из самых волнующих проблем антропологии - о происхождении современного человека.
Ископаемые из отложения Соло не дают покоя эволюционистам уже 60 лет, с самого момента их обнаружения. Проблема в том, что останки имеют явную морфологию Homo erectus, но по геологическому контексту относятся к гораздо более позднему периоду. Такая комбинация никак не устраивает поборников эволюционной теории. Ископаемые останки Homo Erectus с очень поздним периодом залегания найдены и в Австралии, но эволюционисты вышли из положения, произвольно определив их как останки Homo Sapiens
. С Яванским человеком подобное чисто семантическое решение невозможно, поскольку большинство палеонтологов определило их принадлежность к Homo Erectus еще до того, как выяснился их весьма молодой возраст.
В период между 1931 и 1933 гг. группа голландских исследователей обнаружила останки двенадцати человеческих черепов в отложении песчаника полуметровой толщины у реки Соло. Были также найдены кости двух человеческих ног. На площади всего 50х100 м. были обнаружены разрозненные останки более чем 25 тысяч позвоночных животных. Между 1976 и 1980 годами экспедиция из Университета Гаджа Мада , раскапывавшая соседний участок площадью 25х16 м., извлекла еще два человеческих черепа, несколько осколков человеческих тазовых костей, различные человеческие артефакты, а также еще 1200 останков позвоночных животных. Это были не целые человеческие черепа, а фрагменты, в том числе череп без лицевых частей и нижней челюсти, а также крышка черепа.
С самого начала вокруг ископаемых Соло начались споры. Сразу было очевидно, что артефакты эпохи мезолита, найденные вместе с черепами (которые Кеннет Окли назвал "азильской культурой оброботки кости", предполагают датировку в 10 тыс. лет и меньше, ведь австралийские аборигены вплоть до недавнего времени жили на культурном уровне мезолита. 1
Столь позднее датирование представляло опасность для всей теории эволюции человека, и эволюционисты заявили, что человеческие останки и артефакты не связаны между собой и находятся на разных стратиграфических уровнях. Это утверждение, сделанное постфактум, противоречит недвусмысленным свидетельствам очевидцев. Хотя окаменелости действительно найдены до появления многих современных археологических методов, но все-таки раскопки проводились под контролем Голландского геологического обозрения; знаменитый антрополог ван Кенигсвальд [Кoenigswald] много раз бывал на раскопках, видел череп #VI (Нгандонг 7) и череп #VIII (Нгандонг 11) in situ, откопал оба черепа и описал предметы культуры, найденные вместе с ними. 2
История датирования черепов из Соло весьма красочна сама по себе. Поскольку первоначальная находка была сделана до возникновения радиометрического метода, датирование почти целиком основывалось на фауне (ископаемых животных), найденной вместе с черепами. Наиболее ранний предполагаемый возраст ископаемых колебался от 150 тыс. до 100 тыс. лет. Эти оценки были сделаны невзирая на то, что все записи, относящиеся к человеческим черепам и окружавшей их фауне, были утеряны во время II Мировой войны 3 , и "...большинство из 25 тыс. ископаемых первой голландской экспедиции оказались утеряны." 4 Возможность того, что останки предположительного "Homo еrectus"
могут оказаться всего лишь стотысячелетней давности, совершенно не устраивала эволюционистов, и кто-то предположил, что окаменелости животных и людей могут быть разного возраста - человеческие останки гораздо старше. Но неопубликованные фотографии раскопа, сделанные ван Кенигсвальдом, говорят о том, что ископаемые останки людей и позвоночных животных находятся в одном геологическом контексте. 5
Поставив под вопрос возраст животных, найденных в первом раскопе, некоторые эволюционисты стали забавляться "морфологическим датированием", вычисляя увеличение объема мозга с течением времени. В результате возраст черепов из Соло оказался примерно от 463 до 790 тыс. лет. 6 Позже анализ магнитной поляризации дал датировку черепа средним плейстоценом - от 350 до 700 тыс. лет. Во время последних раскопок 1976-1980 годов было обнаружено 1200 ископаемых позвоночных животных. Основываясь на этих окаменелостях, Г.Г.Поуп определил возраст "людей из Соло" примерно в миллион лет. 8 Не удивительно, что последняя датировка, сократившая возраст "человека Нгандонг Соло" до 27 тыс. лет, произвела такое шоковое впечатление. Возраст человеческих останков из Самбунгмакана, в 40 км. выше, оценивался в 1,3 млн. лет; новое исследование дало тот же результат - 27 тыс. лет. 9 Классифицировать останки из Соло оказалось не проще, чем определить их возраст. Когда они были обнаружены, ван Кенигсвальд предположил, что это "тропический неандерталец"
. В 1963 г. Бернард Кемпбелл окрестил их "Homo sapiens soloensis"
("человек разумный из Соло"
). Сента-Лука** в 1980 г. назвал их "Homo ereсtus erectus" ("человек прямоходящий прямоходящий
"), присоединившись к мнению Милфорда Уолпоффа о том, что это не Homo ereсtus. Другие антропологи сочли, что это "древний Homo sapiens
". Из-за явного сходства с яванскими и китайскими "классическими образцами Homo ereсtus" большинство современных исследователей все же причисляет эти останки к "Homo ereсtus
". Однако у ископаемых людей из Соло вместимость черепной коробки больше, и эволюционисты не удержались от искушения назвать их "переходным звеном" между Homo ereсtus и современным человеком. К несчастью, эволюционная теория предполагает, что современный человек появился около 100 тыс. лет назад, и "переходные" ископаемые с возрастом в 27 тыс. лет в эту теорию никак не вписываются.
Положение человеческих черепов и ископаемых животных свидетельствует против гипотезы о том, что их снесло водой вниз по течению. Билс и Хойер пишут: "Все черепа обнаружены лежащими основанием вверх, в отличной сохранности. без всяких следов износа или передвижения". 10 Карлтон Коон подтверждает чуть ли не слово в слово: ""Все черепа лежат основанием вверх, находятся в отличной сохранности. Они не передвигались и не перекатывались." 11 Свишер утверждает, что ископаемые останки млекопитающих выше по течению найдены лишь в 30 км. Далее он говорит, что на раскопе Соло есть "несколько сочлененных позвонков и несколько черепов с присоединенными нижними челюстями...", и что "на черепах гоминидов и негоминидов почти нет следов истирания; сохранились хрупкие отростки." 12 Кроме того, возраст человеческих окаменелостей из Самбунгмакана, в 40 км. выше по течению, не более древний. Все это говорит о том, что ископаемые найдены в их изначальном положении.
С одной стороны, в журнале "Science"
туманно утверждается, что человеческие ископаемые были перенесены потоками воды; но в том же выпуске журнала Энн Гиббонс пишет: "Что касается теории наводнения: группа Свишера подчеркивает, что невозможно представить, как 12 черепов и другие человеческие окаменелости могло принести на один уровень в два разных места (Нгандонг и Самбунгмакан)". 13 Касаясь возможности того, что ископаемые могли быть перенесены водой в более молодые отложения, журнал "Time" пишет: "Свишер отрицает такую возможность, подчеркивая высокую сохранность ископаемых; их хрупкие структуры совершенно не разрушены, то есть, они не попали в воды потопа." 14
Многие современные исследователи согласны с гипотезой ван Кенигсвальда о том, что "люди Соло (Нгандонга)" пали жертвой каннибализма. Он пишет: "большое количество костей всех видов животных лежит в своем естественном состоянии, а от костяков людей сохранился очень странный подбор фрагментов явно не естественного происхождения." 15 У всех черепов без исключения разбиты лицевые кости, и у всех, кроме двух, разбито основание черепа. Ван Кенигсвальд называет их "трофейными черепами", проводя аналогию с современными "охотниками за головами", которые съедают мозг побежденного противника, чтобы овладеть его умом и храбростью. Черепа должны были отмечать границы территории племени. "Даже в наше время некоторые племена Новой Гвинеи отмечают подобным образом границы своих охотничьих угодий. Видимо, они считают, что дух, обитавший в черепе, помогает охранять их территорию от врагов." 16
Раньше эволюционисты довольно успешно отвергали утверждения о позднем происхождении "людей Соло (Нгандонга)" и о том, что они существовали одновременно с современными людьми. Теперь же аргументы в пользу этого стали гораздо убедительней. Крис Стингер из Лондонского Музея Естественной истории, ошибочно считающий неандертальцев отдельным видом, пишет: "Если датировка верна, то мы видим три разных вида, существующих одновременно." 17 Эволюционистам предстоят еще более тяжелые испытания: например, как объяснить то, что многие австралийские окаменелости позднего датирования практически идентичны с "людьми Соло (Нгандонга)"?
Милфорд Уолпофф (Мичиганский университет), говоря о предполагаемой эволюции австралопитека в Homo erectus, заявляет следующее: "...когда оказалось, что "Homo erectus" мог появиться раньше, чем его предполагаемый предок,.. следовало опубликовать опровержение" 18 Уолпофф совершенно прав: именно так следует поступать в палеоантропологии.
За высказыванием Уолпоффа стоит определенный научный принцип. Он гласит следующее: "Эволюционная последовательность ложна, если характерная форма этой последовательности выпадает из собственной эволюционной временной схемы." Именно это и произошло с "человеком из Соло (Нгандонга)".
Библиография:
1 . Kenneth P.Oakley, Frameworks For Dating Fossil Man. Chicago, Aldine Publishing Company, 1964, pp.171-172, 251-252, 314, and chart between pp.170-171. Kenneth P.Oakley, Man The Tool-Maker, sixth edition, Chicago, The University of Chicago Press, 1972, pp.66-70, 80.2 . G.H.R.von Koenigswald, Meeting Prehistoric Man, trans.by Michael Bullock, New York, Harper Publ., 1956, pp.65-79.
3 . W.W.Howells, Homo Erectus - Who, When and Where: A Survey. Yearbook of Physical Anthropology 23, New York, Alan R. Liss, Inc., 1980, p.5.
4 . C.C.Swisher III, W.J. Rink,S.C. Anton, H.P.Schwarcz, G.H.Curtis, A.Suprijo, Widiasmoro, Latest Homo Erectus of Jawa; Potential Contemporaniety with Homo Sapiens in Southeast Asia, Science 274, 13 December 1996, p.1871.
5 . Swisher et al., p.1871.
6 . Howells, p.5, footnote.
7 . Rightmire 14, p.192.
8 . Geoffrey G.Pope, Ngandong (Solo River), Encyclopaedia of Human Evolution and Prehistory, Ian Tattersall, Eric Delson and John Van Couvering, editors, New York, Garland Publishing, 1988, p.383.
9. Swisher et al., p.1871.
10 .Ralph L.Beals and Harry Hoijer, An Introduction to Anthropology, third edition, New York, The Macmillan Company, 1965, p.104.
11 .Carleton S.Coon, The Origin of Races, New York, Alfged A.Knopf, 1962,p.390.
12 .Swisher et al., p.1871.
13 .Ann Gibbons, Homo Erectus in Jawa: A 250,000-Year Anachronism, Science 274, 13 December 1996, p.1841.
14 .Jeffrey Kluger, Not So Extinct After All, Time, December 23, 1996, p.68.
15 .von Koenigswald, 75.
16 .von Koenigswald, 76.
17 .Gibbons 1841.
18 .Millford H.Wolpoff, Paleoanthropology, New York, Alfged A.Knopf, 1980, p.vi.
Перевод Яна Шапиро
Что же такое человек?... У него четыре ноги; две передних служат ему
для хватания, две задних для ходьбы; следовательно, это четвероногое животное...
Д. Дидро (Об объяснении природы, 1754 г.)
4.1.1. Прародина
Каждый исследователь придерживается своей точки зрения на то, по какому варианту шла эволюция предков человека - путем Кладогенез
- возникновение новых видов из маленькой изолированной популяции.");" onmouseout="nd();" href="javascript:void(0);">кладо-
или Анагенез
- одна из основных теоретических форм эволюционного процесса, представляющая собой прямое последовательное развитие без ветвления (чем отличается от кладогенеза - эволюции по типу " onmouseout="nd();" href="javascript:void(0);">анагенеза
. Возможно, на разных этапах она шла по-разному. Ответ зависит также от того, с какого эволюционного момента начинать рассматривать этих предшественников. Мы начнем обзор с миоценовых гоминоидов, с момента разделения линий человекообразных обезьян и человека.
Важнейшим является вопрос о прародине исходной формы человеческого предка.
Азиатская прародина человека. Исторически первой была разносторонне аргументирована гипотеза азиатской прародины. В Пакистане и Индии были сделаны многочисленные находки среднемиоценовых гоминоидов Сивапитеков (Sivapithecus ), живших около 8-12 млн лет назад. До открытий в Африке азиатские обезьяны считались предками гоминид. Однако в Азии нет находок прогрессивных гоминоидов в промежутке времени от 8 до 1,5 млн л., тогда как в Африке существовали австралопитековые. По современным представлениям, азиатские Сивапитеки (Sivapithecus ) - ископаемые понгиды, жившие в Азии около 12-8 млн. лет назад. Важнейшие находки сделаны в Пакистане, Северной Индии и Южном Китае. Внешне и по образу жизни были очень похожи на современных орангутанов. Существовали сивапитеки размером с орангутана и меньше. Судя по строению черепа, сивапитеки были прямыми предками орангутанов. Раньше, до появления африканских находок, сивапитеков считали предками современного человека, но сейчас эта гипотеза отвергнута.");" onmouseout="nd();" href="javascript:void(0);">сивапитеки были предками лишь современных орангутангов.
Африканская прародина человека. Позже азиатских были совершены богатейшие африканские находки ископаемых костей. Из Африки известен целый ряд находок приматов, живших в раннем и среднем миоцене. Древнейшие из этих гоминоидов - проконсулы (Proconsul ) (10-22 млн лет назад) и афропитеки (Afropithecus ) (17-20 млн лет назад), более поздние дриопитеки (Dryopithecus ) (9-16 млн лет назад). Их Морфология (греч. morphe вид, форма) - в биологии учение о вариации строения объекта, его формы и структуры. Аналогичное специальное направление антропологии (собственно морфология) занято изучением вариации строения отдельных органов человеческого тела и их систем, возрастной изменчивости организма человека, его физического развития и конституции.");" onmouseout="nd();" href="javascript:void(0);">морфология наиболее близка к современным понгидам и первым австралопитекам.
Прародина человека в Южной Европе.
Ископаемые гоминоиды жили не только в Азии и Африке, много человекообразных обезьян населяло в миоцене Южную Европу - гориллоподобные Уранопитеки (Ouranopithecus
) и шимпанзеподобные Дриопитеки (Dryopithecus
) (9-10 млн лет назад).
Из трех возможных центров происхождения исходного предка человека наиболее полная связь миоценовых и более поздних гоминоидов прослеживается в Африке: Самбуропитек (Samburupithecus
) (9,5 млн лет назад) - Сахелянтроп (Sahelanthropus
) (6-7 млн лет назад). В Азии и Европе есть довольно поздние миоценовые человекообразные обезьяны, но нет очень древних Гоминиды
- семейство приматов, к которому относят современного человека. По наиболее распространенной схеме включают два подсемейства - австралопитецин и гоминин. Некоторые ученые считают, что к гоминидам можно отнести и человекообразных обезьян.");" onmouseout="nd();" href="javascript:void(0);">гоминид
. Таким образом, Африка с наибольшей вероятностью является прародиной гоминид.
4.1.2. Гоминидная триада
Рамки семейства гоминид зависят от того, какие признаки положены в основу определения места конкретных видов в системе Гоминоиды (Hominoidea ) - высшие обезьяны. Возникли в верхнем эоцене или олигоцене, самые древние формы известны из Египта (оазис Файюм). Первые представители - олигопитециды (Oligopithecidae ) и плиопитециды (Pliopithecidae ) ненадежно отличаются от парапитековых обезьян, их систематика крайне запутана. Иногда все они объединяются в одно семейство плиопитецид в качестве трех подсемейств (Pliopithecinae, Oligopithecinae и Propliopithecinae ). Более прогрессивные формы известны из миоцена, когда гоминоиды достигли своего расцвета. Нижне-среднемиоценовые гоминоиды относятся к Проконсулидам (Proconsulidae ), а средне-поднемиоценовые - к Дриопитецидам (Dryopithecidae ). В миоцене возникли и все три современных семейства: гиббоновые (Hylobatidae ), понгиды (Pongidae ), гоминиды (Hominidae ).");" onmouseout="nd();" href="javascript:void(0);">гоминоидов . Поскольку единственным современным представителем семейства является человек, из его особенностей исторически были выделены три важнейшие системы, считающиеся истинно гоминидными.
- Эти системы были названы гоминидной триадой:
- прямохождение (бипедия);
- кисть, приспособленная к изготовлению орудий;
- высокоразвитый мозг.
1. Прямохождение.
Относительно его происхождения выдвинуто множество гипотез. Двумя важнейшими являются - миоценовое похолодание и трудовая концепция.
Миоценовое похолодание:
в середине и конце миоцена в результате глобального похолодания климата произошло значительное сокращение площадей тропических лесов и увеличение площади саванн. Это могло стать причиной перехода части гоминоидов к наземному образу жизни. Однако, известно, что древнейшие известные прямоходящие приматы жили в тропических лесах.
Трудовая концепция:
согласно широкоизвестной трудовой концепции Ф. Энгельса и ее позднейшим вариантам, возникновение прямохождения тесно связано со специализацией руки обезьяны для трудовой деятельности - переноса предметов, детенышей, манипулирования пищей и изготовления орудий (). В дальнейшем труд привел к возникновению языка и общества. Однако, по современным данным, прямохождение возникло намного раньше изготовления орудий. Прямохождение возникло не менее 6 миллионов лет назад у Orrorin tugenensis
, а древнейшие орудия из Гоны в Эфиопии имеют датировку лишь 2,7 млн лет назад.
Другие гипотезы.
Существуют и иные версии возникновения прямохождения. Оно могло возникнуть для ориентирования в саванне, когда надо было смотреть поверх высокой травы. Также, предки человека могли вставать на задние ноги, чтобы переправляться через водные преграды или пастись на заболоченных лугах, как это делают современные гориллы в Конго.
Согласно концепции К. Оуэна Лавджоя, прямохождение возникло в связи с особой стратегией размножения, поскольку Гоминиды
- семейство приматов, к которому относят современного человека. По наиболее распространенной схеме включают два подсемейства - австралопитецин и гоминин. Некоторые ученые считают, что к гоминидам можно отнести и человекообразных обезьян.");" onmouseout="nd();" href="javascript:void(0);">гоминиды
в течение очень длительного времени выращивают одного, максимум два детеныша (см.: ;). При этом уход за потомством достигает такой сложности, что появляется необходимость освобождения передних конечностей. Переноска беспомощных детенышей и пищи на расстояние становится жизненно важным элементом поведения. Согласно Лавджою, прямохождение возникло еще в тропическом лесу, а в саванны переселились уже двуногие гоминиды (см. Хрестом. 4.1).
Кроме того, экспериментально и на математических моделях доказано, что передвижение на большие расстояния со средней скоростью на двух ногах энергетически более выгодно, чем на четырех.
Вероятнее всего, в эволюции действовала не одна причина, а целый их комплекс.
- Для определения прямохождения у ископаемых приматов ученые пользуются следующими основными признаками:
- положение затылочного отверстия - у прямоходящих оно находится в центре длины основания черепа, открывается вниз. Такое строение известно уже у Sahelanthropus tchadensis и Ardipithecus ramidus ramidus около 4-7 млн лет назад. У четвероногих - в задней части основания черепа, повернуто назад.
- строение таза - у прямоходящих таз широкий и низкий (такое строение известно начиная с Australopithecus afarensis 3,2 млн лет назад), у четвероногих таз узкий, высокий и длинный;
- строение длинных костей ног - у прямоходящих ноги длинные, коленный и голеностопный суставы имеют характерное строение. Такое строение известно, начиная с Orrorin tugenensis 6 млн лет назад. У четвероногих приматов руки длиннее ног.
- строение стопы - у прямоходящих выражен свод (подъем) стопы, пальцы прямые, короткие, большой палец не отведен в сторону, малоподвижен (свод выражен уже у Australopithecus afarensis , но пальцы длинные и изогнутые у всех австралопитеков, у Homo habilis стопа уплощенная, но пальцы прямые, короткие), у четвероногих стопа плоская, пальцы длинные, изогнутые, подвижные. В стопе Australopithecus anamensis большой палец был малоподвижен. В стопе Australopithecus afarensis большой палец противопоставлялся другим, но намного слабее, чем у современных обезьян, своды стопы развиты хорошо, отпечаток ноги был почти как у современного человека. В стопе Australopithecus africanus и Paranthropus robustus большой палец был сильно отведен от других, пальцы были очень подвижные, строение промежуточное между обезьянами и человеком. В стопе Homo habilis большой палец полностью приведен к остальным.
- строение рук - у полностью прямоходящих гоминид руки короткие, не приспособлены к хождению по земле или лазанию по деревьям, фаланги пальцев прямые. Черты приспособления к хождению по земле или лазанию по деревьям есть у австралопитеков Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Paranthropus robustus и даже Homo habilis .
Таким образом, прямохождение возникло более 6 миллионов лет назад, но еще долго отличалось от современного варианта.
Некоторые австралопитеки и Homo habilis
использовали и другие виды передвижения - лазание по деревьям и хождение с опорой на фаланги пальцев рук.
Полностью современным прямохождение стало только около 1,6-1,8 миллиона лет назад.
- 2. Происхождение руки, приспособленной к изготовлению орудий.
Рука, способная изготовлять орудия, отличается от руки обезьяны. Хотя морфологические признаки рабочей руки не являются вполне надежными, однако можно выделить следующий трудовой комплекс:
- Сильное запястье. У австралопитеков, начиная с Australopithecus anamensis , строение запястья промежуточное между обезьянами и человеком. Практически современное строение наблюдается у Homo habilis 1,8 млн лет назад.
- Противопоставление большого пальца кисти. Признак известен уже 3,2 млн лет назад у Australopithecus afarensis и Australopithecus africanus. Он был полностью развито у Paranthropus robustus и Homo habilis 1,8 млн лет назад. Наконец, было своеобразным или ограниченным у неандертальцев Европы около 40-100 тыс. лет назад.
- Широкие конечные фаланги пальцев. Очень широкие фаланги были у Paranthropus robustus, Homo habilis и всех более поздних гоминид.
- Прикрепление мышц, двигающих пальцы почти современного типа отмечено у Paranthropus robustus и Homo habilis , но у них есть и примитивные черты.
Кости кисти у древнейших прямоходящих гоминоидов (Orrorin tugenensis, Ardipithecus ramidus kadabba, Australopithecus anamensis и Australopithecus afarensis
) имеют смесь признаков человекообразных обезьян и человека. Скорее всего, эти виды могли использовать предметы как орудия, но не изготовлять их. Первые изготовители настоящих орудий - Homo habilis
. Вероятно, орудия изготовляли и южноафриканские массивные австралопитеки Paranthropus robustus
(рис. 4.1).
Итак, трудовая кисть в целом сформировалась около 1,8 млн лет назад
.
- 3. Высокоразвитый мозг.
Мозг современного человека сильно отличается от мозга человекообразных обезьян по размерам, форме, строению и функциям, однако среди ископаемых форм можно найти множество переходных вариантов. Типичные признаки мозга человека следующие:
- Большие общие размеры мозга. У австралопитеков размер мозга был как у современных шимпанзе. Бурный рост размеров происходил у Homo habilis около 2,5-1,8 млн лет назад, а у позднейших Гоминиды - семейство приматов, к которому относят современного человека. По наиболее распространенной схеме включают два подсемейства - австралопитецин и гоминин. Некоторые ученые считают, что к гоминидам можно отнести и человекообразных обезьян.");" onmouseout="nd();" href="javascript:void(0);">гоминид наблюдается плавное увеличение до современных значений.
- Специфические поля мозга - зоны Брока и Вернике и другие поля начали развиваться у Homo habilis и архантропов, но полностью современного вида достигли, видимо, только у современного человека.
- Строение долей мозга. У человека значительно развиты нижняя теменная и лобная доли, острый угол схождения височной и лобной долей, височная доля широкая и округлая спереди, затылочная доля относительно небольшая, нависает над мозжечком. У австралопитеков строение и размеры мозга были такие же, как и у человекообразных обезьян. Строение мозга Homo habilis было примитивным, но сам мозг имел большие размеры. Для позднейших гоминид характерно плавное приближение к современному строению мозга.
Гоминидный тип мозга возник около 1,8 млн лет назад или несколько раньше, причем увеличение размеров предшествовало его усложнению (рис. 4.2). В дальнейшем мозг активно эволюционировал, а полностью современный мозг сформировался только около 25 тыс. лет назад
(см. Хрестом. 4.2).
К гоминидной триаде можно добавить такой надежный признак, как маленькие клыки
. Современные и ископаемые человекообразные обезьяны имеют крупные клыки, выступающие за линию других зубов. Древнейшие гоминиды и австралопитеки, напротив, обладали небольшими клыками. Насколько известно на сегодняшний день, все четвероногие гоминоиды - обладатели больших клыков, а прямоходящие - маленьких (рис. 4.3).
Мы видим, что древнейшие гоминиды отличались от человека по многим биологическим характеристикам, и из гоминидной триады единственным надежным признаком семейства является прямохождение.
Итак: гоминиды (семейство Hominidae) - это прямоходящие приматы с маленькими клыками.
4.1.3. Наши предшественники - наши предки?
В семейство гоминид включают современного человека и его непосредственных предшественников. Обычно древнейшей границей этой группы условно считают момент разделения общей эволюционной линии на ветви, ведущие к современным человекообразным обезьянам и современному человеку.
- Наиболее принятым в современной науке является выделение в семействе гоминид (Hominidae
) двух подсемейств:
- австралопитецины (Australopithecinae ) - гоминиды с множеством типичных понгидных черт;
- гомиинны (Homininae ) - гоминиды без понгидных черт.
1. Австралопитецины
(Australopithecinae
). Древнейшими гоминидами обычно считают австралопитековых.
Australopithecinae
) - группа приматов, по строению и поведению промежуточная между обезьянами и людьми или, точнее, понгидами и гоминидами. Прямоходящие приматы с обезьяньей головой, маленьким мозгом и небольшими клыками. Наиболее подходят под определине " onmouseout="nd();" href="javascript:void(0);">Австралопитековые
были очень своеобразной группой. Кем они были - двуногими обезьянами или людьми с обезьяньей головой? И как относиться к такому сочетанию признаков?
Появились австралопитековые около 6-7 млн лет назад, а последние из них вымерли только около 900 тыс. лет назад, во время существования намного более прогрессивных форм. Насколько известно, австралопитековые никогда не покидали пределов Африки, хотя некоторые находки, сделанные на острове Ява, иногда относят к этой группе.
Сложность положения австралопитековых среди приматов заключается в том, что в их строении мозаично сочетаются признаки, характерные и для современных человекообразных обезьян, и для человека.
Череп австралопитеков похож на череп шимпанзе. Характерны большие челюсти, массивные костные гребни для прикрепления жевательной мускулатуры, маленький мозг и большое уплощенное лицо. Зубы австралопитеков были очень большие, но клыки короткие, а детали строения зубов больше похожи на человеческие, чем обезьяньи.
В строении скелета австралопитеков характерны широкий низкий таз, относительно длинные ноги и короткие руки, хватательная кисть и нехватательная стопа, вертикальный позвоночник. Такое строение уже почти человеческое, отличия заключаются лишь в деталях строения и в маленьких размерах.
Рост австралопитеков колебался от метра до полутора. Характерно, что размер мозга был около 350-550 см 3 , то есть как у современных горилл и шимпанзе. Для сравнения, мозг современного человека имеет объем около 1200-1500 см 3 . Строение мозга австралопитеков также было весьма примитивно и мало отличалось от шимпанзоидного.
Образ жизни австралопитеков, видимо, был непохож на известный у современных приматов. Они жили в тропических лесах и саваннах, питались преимущественно растениями. Впрочем, поздние австралопитеки охотились на антилоп или отнимали добычу у крупных хищников - львов и гиен.
Австралопитеки жили группами в несколько особей и, видимо, постоянно кочевали по просторам Африки в поисках пропитания. Орудия австралопитеки вряд ли умели изготовлять, хотя использовали наверняка. Их руки были весьма похожи на человеческие, но пальцы были сильнее изогнуты и более узкие. Как уже упоминалось, древнейшие орудия известны из слоев в Эфиопии, датированных 2,7 млн лет назад, то есть спустя 4 млн лет после появления австралопитеков. В Южной Африке австралопитеки или их непосредственные потомки около 2-1,5 млн лет назад использовали костяные обломки для вылавливания термитов из термитников.
- Австралопитеки
(Австралопитековые, или австралопитецины, Australopithecinae
) - группа приматов, по строению и поведению промежуточная между обезьянами и людьми или, точнее, понгидами и гоминидами. Прямоходящие приматы с обезьяньей головой, маленьким мозгом и небольшими клыками. Наиболее подходят под определине " onmouseout="nd();" href="javascript:void(0);">Австралопитеков
можно разделить на три основных группы, в каждой из которых выделяется несколько видов:
- Ранние австралопитеки - существовали с 7 до 4 млн лет назад, обладали наиболее примитивным строением. Выделяют несколько родов и видов ранних австралопитеков.
- Грацильные австралопитеки - существовали с 4 до 2,5 млн лет назад, имели сравнительно небольшие размеры и умеренные пропорции. Обычно выделяют один род Australopithecus c несколькими видами.
- Массивные австралопитеки - существовали с 2,5 до 1 млн лет назад, были очень массивно сложенными специализированными формами с крайне развитыми челюстями, маленькими передними и огромными задними зубами. Массивные австралопитеки выделяются в самостоятельный род Paranthropus с тремя видами. Рассмотрим более подробно каждую из них.
Остатки древнейших приматов, которых можно отнести к Ранние австралопитеки
- наиболее древние представители австралопитеков, жившие с 7 до 4 миллионов лет назад. Наиболее подходят на роль пресловутого " onmouseout="nd();" href="javascript:void(0);">ранним австралопитекам
, были найдены в Республике Чад в Торос-Меналла и названы Сахелянтроп
(Sahelanthropus tchadensis
) - самый ранний гоминид, относимый к кругу ранних австралопитековых. Описан в 2002 г. по находке в Республике Чад. Датировка по фауне около 6-7 млн. лет назад. Среди находок - нижняя челюсть и целый череп, который получил популярное название " .="" onmouseout="nd();" href="javascript:void(0);">Сахелянтроп
(Sahelanthropus tchadensis
). Целый череп получил популярное название "Тумай". Датировка находок около 6-7 млн лет назад. Более многочисленные находки в Кении в Туген Хиллс имеют датировку 6 млн. лет назад. Они получили название Оррорин
(Orrorin tugenensis
) - древнейший гоминид, относимый к кругу ранних австралопитековых. Описан в 2001 г. Останки (кости конечностей и зубы) обнаружены в Кении в местонахождении Туген Хиллс и имеют датировку 6 млн. лет назад. Клыки Орроринов были сравнительно с друними гоминидами большие, но меньше, чем у шимпанзе. Судя по строению бедренных костей, Оррорины были прямоходящими, ростом примерно 1,4 м. Однако фаланги пальцев у них изогнутые, что можно расценивать и как признак лазания по деревьям.");" onmouseout="nd();" href="javascript:void(0);">Оррорин
(Orrorin tugenensis
). В Эфиопии в двух местонахождениях - Алайла и Арамис - были найдены многочисленные костные останки, названные Ардипитек
(Ardipithecus ramidus
) - одни из самых ранних австралопитеков, от которых известны достаточно многочисленные останки. Найдены в Эфиопии. Более древний подвид Ardipithecus ramidus kadabba
обнаружен в местонахождении Алайла и датирован около 5,5 млн. лет назад. Описан в 2001 г. Более молодой подвид Ardipithecus ramidus ramidus
найден в местонахождении Арамис и датирован 4,4 млн. лет назад. Описан в 1994 и 1995 г. Судя по строению основания черепа, Ардипитеки были прямоходящими существами ростом около 1,2 м и весом около 40 кг. Зубы сочетают признаки шимпанзе и человека, то же можно сказать про кости стопы, руки и кисти. Высказано предположение, что Ардипитеки являются предками современных шимпанзе.");" onmouseout="nd();" href="javascript:void(0);">Ардипитек
(Ardipithecus ramidus kadabba
) (около 5,5 млн лет назад) и Ardipithecus ramidus ramidus
(4,4 млн. лет назад). Находки в двух местонахождениях в Кении - Канапои и Аллия Бей - получили название Австралопитек анамский
(Australopithecus anamensis
) - найден в Кении в местонахождениях Канапои и Аллия Бей, описан в 1995 г. Датируется временем около 4 млн. лет назад. Известен по находкам челюстей, зубов и нескольких костей скелета. Все останки сочетают понгидные и гоминидные признаки примерно в равной пропорции. Некоторые черты челюстей и зубов могут быть охарактеризованы как специализированные, что, вероятно, исключает Австралопитека анамского из числа прямых предков современного человека. Особенно примитивна локтевая кость, имеющая черты, свидетельствующие о передвижении с опорой на фаланги пальцев, как у современных шимпанзе и горилл. Прямохождение недоказано.");" onmouseout="nd();" href="javascript:void(0);">Австралопитек анамский
(Australopithecus anamensis
). Они датируются временем 4 млн. лет назад (рис. 4.4).
Рост их ненамного превышал один метр. Размеры мозга были такими же, как у шимпанзе. Обитали ранние австралопитеки в лесистых или даже болотистых местах, а также в лесостепях. Очевидно, именно эти существа наиболее подходят на роль пресловутого "промежуточного звена" между обезьяной и человеком. Мы практически ничего не знаем про их образ жизни, однако с каждым годом количество находок растет, а знания об окружающей среде того далекого времени ширятся (см. Доп. материал 4.1).
О ранних австралопитеках известно не очень много (табл. 4.1). Судя по черепу Сахелянтропа, бедренным костям Оррорина, фрагментам черепа, костям конечностей и остаткам таза Ардипитеков, ранние австралопитеки были уже прямоходящими приматами. Однако, судя по костям рук Оррорина и Австралопитека анамского, они сохраняли способность лазания по деревьям или даже были четвероногими существами, опиравшимися на фаланги пальцев рук, подобно современным шимпанзе и гориллам. Строение зубов ранних австралопитеков промежуточно между обезьянами и человеком. Возможно даже, Сахелянтропы были родственниками горилл, Ардипитеки - непосредственными предками современных шимпанзе, а анамские австралопитеки вымерли, не оставив потомков.
| Название (первое описание) | Синонимы | Регион | Время, млн. л. |
|---|---|---|---|
| Sahelanthropus tchadensis (Brunet et al., 2002) | - | Чад | 6-7 |
| Orrorin tugenensis (Senut et al., 2001) | - | Кения | 5,6-6,2 |
| Ardipithecus ramidus (White et al., 1995) | =Homo antiquus praegens
(Ferguson, 1989) =Australopithecus praegens |
Эфиопия | 3,9-5,5 |
| Australopithecus anamensis (Leakey et al., 1995) | - | Кения | 3,9-4,2 |
Приблизительно с 4 млн лет назад палеонтологическая летопись становится достаточно полной и последовательной.
В Кении, Танзании и Эфиопии во множестве местонахождений открыты окаменелости Грацильные австралопитеки
- группа австралопитеков (Australopithecus
), существовавшая от 4 до 2,5 миллиона лет назад. Имели сравнительно небольшие размеры и умеренные пропорции. Были потомками ранних австралопитеков и предками массивных австралопитеков (или парантропов) и ">Homo".");" onmouseout=nd(); href="javascript:void(0);">грацильных австралопитеков
, названные Австралопитек афарский
(Australopithecus afarensis
) - восточноафриканский вид грацильных австралопитеков, описанный в 1978 г. Существовал от 4 до 2,5 млн. лет назад. Наиболее известны находки из местности Хадар в пустыне Афар, в том числе скелет, получивший прозвище Люси. Судя по костям конечностей и таза, Австралопитеки афарские были, несомненно, прямоходящими существами ростом около 1-1,5 м и весом 30-60 кг. Вместе с тем, фаланги пальцев кисти у них изогнуты, так что не исключено, что какое-то время эти существа проводили на деревьях, например, ночью. В противоположность прогрессивному строению тела, череп примитивен. Мозг маленький, около 350-500 см 3 . Челюсти большие, сильно выступающие вперед, как у шимпанзе, а на черепе развиты рельефные костные гребни для прикрепления мощных жевательных мышц. Зубы крупные. Афарские или похожие австралопитеки были, видимо, предками всех более поздних гоминид.");" onmouseout="nd();" href="javascript:void(0);">Австралопитек афарский
(Australopithecus afarensis
). Этот вид существовал приблизительно от 4 до 2,5 млн лет назад. Наиболее известны находки из местности Хадар в пустыне Афар, в том числе скелет, получивший прозвище Люси. Также, в Танзании обнаружены окаменевшие следы прямоходящих существ в тех же слоях, в которых найдены останки афарских австралопитеков (рис. 4.5).
Кроме афарских австралопитеков, в Восточной и Северной Африке в промежутке времени 3-3,5 млн лет назад, вероятно, обитали и другие виды. В Кении в Ломекви найден череп и другие окаменелости, описанные как Кениантроп
(Kenyanthropus platyops
) - череп, челюсть и зубы, найденные в Кении в местонахождении Ломекви, отнесены в 2001 г. к особому роду австралопитеков. Они имеют датировку 3,2-3,5 млн. лет назад. Особенностью являются очень плоское лицо, маленькие зубы и др. частные детали морфологии. Некоторые антропологи склонны рассматривать Кениантропа как предка Человека рудольфского.");" onmouseout="nd();" href="javascript:void(0);">Кениантроп
(Kenyanthropus platyops
).
В Республике Чад, в Коро-Торо (Восточная Африка), обнаружен единственный фрагмент челюсти, описанный как Австралопитек бахр-эль-газальский
() - фрагмент челюсти, обнаруженный в Республике Чад, в Коро-Торо, и датированный по фауне 3,0-3,5 млн. лет назад, описанный в 1996 г. как самостоятельный вид. Многие антропологи считают его австралопитеком афарским. Важность находки заключается в ее географическом положении: большинство австралопитеков найдено в Восточной и Южной Африке, а Австралопитек бахр-эль-газальский - в Северной, что свидетельствует о очень широком распространении группы.");" onmouseout="nd();" href="javascript:void(0);">Австралопитек бахр-эль-газальский
(Australopithecus bahrelghazali
) (рис. 4.6).
В Южной Африке, в ряде местонахождений - Таунг, Стеркфонтейн и Макапансгат - обнаружены многочисленные окаменелости, известные под названием Австралопитек африканский
(Australopithecus africanus
) - первый из описанных видов австралопитеков (т.н. " .="" onmouseout="nd();" href="javascript:void(0);">Австралопитек африканский
(Australopithecus africanus
). К этому виду принадлежала первая находка австралопитека - череп детеныша, известного как Бэби из Таунга. Африканские австралопитеки жили с 3,5 до 2,4 млн. лет назад (рис. 4.7).
Самый поздний грацильный австралопитек - с датировкой около 2,5 млн. лет назад - обнаружен в Эфиопии в Боури и назван Австралопитек гари
(Australopithecus garhi
) - самый поздний грацильный австралопитек с датировкой около 2,5 млн. лет назад. Обнаружен в Эфиопии (Боури). Описан в 1999 г. Представлен череп, зубы, кости конечностей. Рост 1,2-1,5 м. Объем мозга около 440 см 3 . Характерной особенностью являются очень большие передние зубы. Вместе с останками А. гари были найдены простейшие каменные орудия и кости антилоп со следами надрезов. Возможно, этот вид был прямым предком линии ведущей к современному человеку.");" onmouseout="nd();" href="javascript:void(0);">Австралопитек гари
(Australopithecus garhi
) (рис. 4.8).
От грацильных австралопитеков известны все части скелета от множества особей, поэтому реконструкции их внешнего облика и образа жизни весьма достоверны (табл. 4.2).
| Название (первое описание) | Синонимы | Регион | Время, млн. л. |
|---|---|---|---|
| Australopithecus afarensis | =Australopithecus africanus tanzaniensis (Tobaios) =Australopithecus aethiopicus (Tobias, 1980) =Homo antiquus (Ferguson, 1984) =Meganthropus africanus (Weinert, 1950) =Praeanthropus africanus (Hennig, 1948; sensu Senuyrek, 1955) =Paranthropus africanus (Weinert, 1950; sensu Olson, 1981) |
Кения, Танзания, Эфиопия | 3,0-3,9-4,4 |
| Kenyanthropus platyops (Leakey et al., 2001) | - | Кения, Эфиопия | 3,2-3,5 |
| Australopithecus bahrelghazali (Brunet et al., 1996) | - | Чад | 3,0-3,5 |
| Australopithecus africanus | =Australopithecus prometheus
(Dart, 1948) = (Broom, 1936) =Homo transvaalensis (Mayr, 1950) |
Южная и Восточная Африка | 2,4-3,5 |
| Australopithecus garhi (Asfaw et al., 1999) | - | Эфиопия | 2,5 |
Грацильные австралопитеки были прямоходящими существами около 1-1,5 метра ростом. Походка их несколько отличалась от походки человека. Видимо, австралопитеки шагали более короткими шажками, а тазобедренный сустав при ходьбе разгибался не полностью. Вместе с достаточно современным строением ног и таза, руки австралопитеков были несколько удлинены, а пальцы приспособлены для лазания по деревьям, но эти признаки могут быть лишь наследством от древних предков.
Днем австралопитеки кочевали по саванне или лесам, по берегам рек и озер, а вечером забирались на деревья, как это делают современные шимпанзе. Австралопитеки жили небольшими стадами или семьями и были способны перемещаться на довольно большие расстояния. Питались они в основном растительной пищей, а орудий труда обычно не изготовляли, хотя недалеко от костей Австралопитека гари ученые нашли каменные орудия и раздробленные ими кости антилоп. Также, для южноафриканских австралопитеков была выдвинута гипотеза остеодонтокератической (дословно - "костно-зубо-роговой") культуры. Предполагалось, что австралопитеки использовали кости, рога и зубы животных в качестве орудий. Позднейшие исследования показали, что большинство следов износа появилось на этих костях в результате погрызов гиен и других хищников.
Как и ранние представители рода, Грацильные австралопитеки
- группа австралопитеков (Australopithecus
), существовавшая от 4 до 2,5 миллиона лет назад. Имели сравнительно небольшие размеры и умеренные пропорции. Были потомками ранних австралопитеков и предками массивных австралопитеков (или парантропов) и ">Homo".");" onmouseout=nd(); href="javascript:void(0);">грацильные австралопитеки
имели обезьяноподобный череп, сочетавшийся с почти современным остальным скелетом. Мозг австралопитеков был похож на обезьяний как по размерам, так и по форме. Однако отношение массы мозга к массе тела у этих приматов было промежуточным между небольшим обезьяньим и очень большим человеческим (см. Доп. материал 4.2).
Наиболее вероятными предками человеческой линии эволюции являются афарские австралопитеки
.
Древнейшие каменные орудия известны из нескольких местонахождений в Эфиопии - Гона, Шунгура, Хадар - и имеют датировки 2,5-2,7 млн. лет назад. Тогда же возникли новые виды гоминид, обладавшие большим мозгом и относимые уже к роду Homo. Однако была еще одна группа поздних австралопитеков, уклонившаяся от линии, ведущей к человеку - Массивные австралопитеки
(Парантропы, Paranthropus
) - поздняя группа австралопитеков, существовавшая с 2,5 до 1 миллиона лет назад. Отличаются крайне развитыми челюстями и огромными коренными зубами. Специализированные вегетарианцы, хотя сохранили двуногость и, возможно, умели изготовлять простейшие орудия труда.");" onmouseout="nd();" href="javascript:void(0);">массивные австралопитеки
(табл. 4.3).
| Название (первое описание) | Синонимы | Регион | Время, млн. л. |
|---|---|---|---|
| Paranthropus aethiopicus | =Australopithecus walkeri
(Ferguson, 1989) = (Arambourg et Coppens, 1968) |
Кения, Эфиопия | 2,3-2,6 |
| Paranthropus boisei (Leakey, 1959) | =Zijanthropus boisei (Leakey, 1959) | Восточная Африка | 1,1-2,3-2,5 |
| Paranthropus robustus | =Paranthropus crassidens (Broom, 1948) | Южная Африка | 0,9-2,5 |
Древнейшие массивные австралопитеки известны из Кении и Эфиопии - Локалеи и Омо. Они имеют датировки около 2,5 млн лет назад и названы Парантроп эфиопский
(Paranthropus aethiopicus
) - древнейший массивный австралопитек. Нижние челюсти, череп и зубы найдены в Кении и Эфиопии в местонахождениях Локалеи и Омо. Они имеют датировки около 2,5 млн. лет назад. Описан в 1968 г. От грацильных австралопитеков отличается огромными размерами задних зубов - премоляров и моляров и чрезвычайной массивностью челюстей. От поздних массивных австралопитеков отличается сильно выступающими вперед челюстями и меньшими размерами зубов. Характерны мощные гребни для прикрепления жевательных мышц. Объем мозга очень маленький - всего 410 см 3 . Масса тела, вероятно, около 50-70 кг.");" onmouseout="nd();" href="javascript:void(0);">Парантроп эфиопский
(Paranthropus aethiopicus
) (рис. 4.9).
Более поздние массивные австралопитеки из Восточной Африки - Олдувай, Кооби-Фора - с датировками от 2,5 до 1 млн лет назад описаны как Парантроп Бойса
(Paranthropus boisei
) - поздние восточноафриканские массивные австралопитеки обнаружены в местонахождениях Олдувай, Кооби-Фора, Локалеи и в ряде других. Они датированы от 2,5 до 1 млн. лет назад. Вид описан в 1959 г. по черепу из Олдувая. Наиболее массивный из всех австралопитеков: характерны огромные моляры, тяжелые челюсти, которые, однако, не выступают вперед, большие костные гребни, служившие для прикрепления жевательных мышц. Общая массивность сложения была, видимо, также очень большой - при росте 1,2-1,6 м парантропы Бойса весили от 40 до 90 кг. Парантропы Бойса были полностью прямоходящими. Объем мозга 400-500 см 3 . Важно, что вместе с Парантропами Бойса найдены каменные орудия.");" onmouseout="nd();" href="javascript:void(0);">Парантроп Бойса
(Paranthropus boisei
) (рис. 4.10).
В Южной Африке - Сварткранс, Кромдраай, Дримолен Кэйв - известны Парантропы массивные
(Paranthropus robustus
) - второй описанный вид австралопитеков, получивший название в 1939 г. Несколько сотен их останков обнаружены в Южной Африке в местонахождениях Сварткранс, Кромдраай, Дримолен Кэйв и некоторых других. Парантропы массивные были самым поздним видом австралопитеков. Появились они около 2,5 млн. лет назад, а вымерли только около 900 тыс. лет назад, когда на той же территории жили уже первые люди. Характерны огромные зубы, большие гребни для прикрепления жевательных мышц, тяжелые челюсти. Рост 1,0-1,6 м, вес 40-80 кг. Объем мозга 470-500 см 3 . Парантропы массивные были полностью прямоходящими, а их кисть была приспособлена для изготовления и применения орудий. Каменные и костяные орудия также были найдены, однако они могли быть сделаны и первыми людьми.");" onmouseout="nd();" href="javascript:void(0);">Парантропы массивные
(Paranthropus robustus
). Массивные парантропы были вторым открытым видом австралопитеков (рис. 4.11).
При рассмотрении черепа парантропов бросаются в глаза огромные челюсти и большие костные гребни, служившие для прикрепления жевательных мышц. Максимального развития челюстной аппарат достиг у восточноафриканских парантропов. Первый открытый череп этого вида из-за размеров зубов даже получил прозвище "Щелкунчик".
Прантропы были крупными - до 70 кг весом - специализированными растительноядными существами, жившими по берегам рек и озер в густых зарослях. Образ жизни их в чем-то напоминал образ жизни современных горилл. Тем не менее, они сохранили двуногую походку и даже, возможно, умели изготовлять орудия труда. В слоях с парантропами найдены каменные орудия и костяные обломки, которыми гоминиды разрывали термитники. Также и кисть этих приматов была приспособлена для изготовления и применения орудий (рис. 4.12).
Парантропы "сделали ставку" на размеры и растительноядность. Это привело их к Экологическая специализация
- изменение формы, функции или поведения организма, в результате адаптации к данным условиям среды. Специализация может зайти настолько далеко, что попыляция (вид) перестает быть пластичной по отношению к дальнейшим изменениям среды.");" onmouseout="nd();" href="javascript:void(0);">экологической специализации
и вымиранию. Однако в одних слоях с парантропами обнаружены останки и первых представителей гоминин - так называемых "ранних Homo" - более прогрессивных гоминид с крупным мозгом (см. Доп. материал 4.3).
4.1.4. Первые люди - наши предки
3. Возникновение современного человека (сапиентация).
Взгляды на происхождение современного человека - сапиентацию - с развитием науки значительно менялись. В настоящее время существуют несколько альтернативных взглядов на эту проблему. Все они строго аргументируются, но ни одна не может победить другую.
Сапиентация - процесс возникновения современного вида человека Homo sapiens sapiens, заключающийся как в биологической перестройке - увеличении мозга, округлении черепа, уменьшении размеров лица, появлении подбородочного выступа, - так и в социокультурных новшествах - возникновении искусства, символического поведения, техническом прогрессе, развитии языков.
Прежде всего, существует несколько мнений о том, кого же стоит считать современным человеком? От ответа зависит следующий вопрос - к какому времени обратить взор в поисках прародины? У авторов начала XX в. вопрос о возникновении человека был вопросом о возникновении рас. Затем, с новыми находками и датировками хронологический момент возникновения "первого современного человека" постоянно отодвигался, тогда как момент разделения рас оставался на прежнем месте. В настоящее время возникновение современного вида и появление современных рас превратились в две самостоятельные проблемы и обычно рассматриваются отдельно.
Где же мы находим первые следы первых людей, неотличимых от нас? В нескольких африканских местонахождениях с датировками от 200 до 100 тыс. лет назад обнаружены кости людей, не имевших сильно выступавшего затылка, большого надбровного валика и обладавших при этом очень большим мозгом и выступающим подбородком. Похожие находки были сделаны на Ближнем Востоке - в пещерах Схул и Кафзех. Со времени около 40 тыс. лет назад люди совершенно современного облика, только несколько более массивные, чем мы, - Неоантропы
- стадия человеческой эволюции, включающая и современного человека. Соответствуют ситематическому названию Homo sapiens sapiens
. Во времени следуют за палеоантропами, то-есть, согласно стадиальной теории, появились около 40 тысяч лет назад. Отличаются большим мозгом, наличием выступающего подбородка, очень сложным поведением, включающем язык и искусство. Культура неоантропов времени от 40 до 10 тысяч лет назад называется верхний палеолит. По современным данным, люди современного облика жили уже около 100 тысяч лет назад и были носителями культуры мустье.");" onmouseout="nd();" href="javascript:void(0);">неоантропы
- известны практически со всей территории Ойкумена
- вся населенная человеком территория Земли (видовой ареал современного человека).");" onmouseout="nd();" href="javascript:void(0);">ойкумены
- из Африки, Европы, Азии и Австралии. Только Америка, возможно, была заселена несколько позже.
Население Европы, относящееся к современному виду, жившее в эпоху Верхний палеолит
- комплекс археологических культур, появившийся около 40 и перешедший в мезолит около 10 тысяч лет назад. Прогрессивная и экономная техника изготовления орудий на пластинах позволила резко расширить набор каменных инструментов. Во время верхнего палеолита появляется также обработка кости и искусство. Носителями верхнего палоеолита были первые неоантропы и, возможно, прогрессивные палеоантропы.");" onmouseout="nd();" href="javascript:void(0);">верхнего палеолита
- от 40 до 10 тыс. лет назад - называется Кроманьонцы
- термин неоднозначен: 1) в узком смысле кроманьонцы - это люди, обнаруженные в гроте Кро-Маньон (Франция) и жившие около 30 тыс. лет назад; 2) в более широком смысле это все население Европы времени верхнего палеолита - от 40 до 10 тыс. лет назад; 3) наконец, в наиболее широком понимании кроманьонцами называют всех людей земного шара времени верхнего палеолита. Важнейшие ископаемые находки: в Африке
в Европе
в Азии
в Австралии
в Америке
- Лос Анжелес (23,6 тыс. лет), Лагуна Бич (17 тыс. лет), Монте-Верде (13 тыс. лет), Лагоа-Санта (11,5 тыс. лет), Тепекспан (11 тыс. лет). Почти в каждом из этих местонахождениях обнаружены останки нескольких индивидов, часто разного возраста и пола, в большинстве случаев использован " onmouseout="nd();" href="javascript:void(0);">кроманьонцами
. Легко заметить, что кроманьонцы в Европе 5 тыс. лет подряд соседствовали с Неандертальцы
- часть неандертальцев жила в суровых условиях ледникового периода, что повлекло сильную биологическую и культурную специализацию.");" onmouseout="nd();" href="javascript:void(0);">неандертальцами
. Различались они между собой не только по чертам физического строения. Кроманьонцы обладали намного более совершенной культурой. Неизмеримо выросла техника изготовления орудий. Они стали изготавливаться с пластин - специально подготовленных заготовок, что позволило выделывать куда более изящные орудия, чем мустьерские остроконечники. Кроманьонцы широко использовали для изготовления орудий также кости животных. Выросло техническое оснащение людей - появились лук и стрелы (рис. 4.24).
Существеннейшим явлением является расцвет верхнепалеолитического искусства. В пещерах Франции, Испании и Италии сохранились превосходные образцы наскальной живописи, в слоях стоянок от Бретани до Байкала обнаружены статуэтки людей и животных, сделанные из костей и известняка. Рукоятки ножей и копьеметалок украшались затейливой резьбой. Одежда украшалась бусами и раскрашивалась охрой (рис. 4.25) (см. Хрестом. 4.4).
Искусство имело в то время магическое значение. Изображения животных сопровождаются знаками стрел и копий, призванных облегчить предстоящую охоту. Судя по следам подростков в глине перед пещерными росписями, здесь проводилось и посвящение в охотники. Конечно, мы можем лишь предполагать истинное значение этих следов духовной жизни наших предков, но ее богатство и принципиальное сходство психики людей того времени с нашей несомненно.
Поселения людей Верхний палеолит
- комплекс археологических культур, появившийся около 40 и перешедший в мезолит около 10 тысяч лет назад. Прогрессивная и экономная техника изготовления орудий на пластинах позволила резко расширить набор каменных инструментов. Во время верхнего палеолита появляется также обработка кости и искусство. Носителями верхнего палоеолита были первые неоантропы и, возможно, прогрессивные палеоантропы.");" onmouseout="nd();" href="javascript:void(0);">верхнего палеолита
обычно представляли собой регулярно посещаемые охотничьи Стоянка
- в археологии место кратковременного (однократного, сезонного) проживания группы людей.");" onmouseout="nd();" href="javascript:void(0);">стоянки
. Здесь строились жилища, шла жизнь общества, исполнялись ритуалы, хоронили умерших. Ритуальная практика достигла своего расцвета. В могилу с усопшим кроманьонцы клали орудия, копья, каменные ножи, многочисленные украшения. При этом погребение засыпалось красной охрой, сверху иногда покрывалось костями мамонтов. Очевидно, в это время возникают представления о загробном мире.
В эпоху верхнего палеолита человек приручил волка, превратив его в собаку. Так человек начал сам активно влиять на процесс видообразования у животных (явление т.н. искусственного отбора).
О населении Африки и Азии времени верхнего палеолита известно гораздо меньше, чем о населении Европы. Однако принципиально они были схожи как в биологическом, так и в культурном плане.
Откуда же появился столь понятный нам мир, как он совмещался с совершенно иным миром неандертальцев? Некоторые биологические особенности древнейших верхнепалеолитических людей позволяют предположить, что они пришли в Европу из тропических областей. Длинные конечности, высокий рост, вытянутые пропорции тела, крупные челюсти, вытянутая мозговая коробка схожи у современных тропических популяций и кроманьонцев. Последние отличаются только большими размерами костей, сильным рельефом черепа, более грубыми чертами.
Но, если Кроманьонцы
- термин неоднозначен: 1) в узком смысле кроманьонцы - это люди, обнаруженные в гроте Кро-Маньон (Франция) и жившие около 30 тыс. лет назад; 2) в более широком смысле это все население Европы времени верхнего палеолита - от 40 до 10 тыс. лет назад; 3) наконец, в наиболее широком понимании кроманьонцами называют всех людей земного шара времени верхнего палеолита. Важнейшие ископаемые находки: в Африке
- Кейп-Флетс, Фиш Хук (около 35 тыс. лет), Назлет Хатер (33 тыс. лет); в Европе
- Комб Капелль (около 40 тыс. лет), Младеч (от 38 до 20 тыс. лет), Кро-Маньон (около 30 тыс. лет), Сунгирь и Пшедмости (около 26 тыс. лет), Дольни Вестонице и Гримальди (около 25 тыс. лет); в Азии
- Ниа (40 тыс. лет), Дуньдяньянь (30-10 тыс. лет), Аитапе (19 тыс. лет), Чжоукоудянь (10 тыс. лет); в Австралии
- Мунго (19-14 тыс. лет), Кохуна, Кейлор и Талгай (все около 12 тыс. лет), Коу Свэмп (около 10 тыс. лет); в Америке
- Лос Анжелес (23,6 тыс. лет), Лагуна Бич (17 тыс. лет), Монте-Верде (13 тыс. лет), Лагоа-Санта (11,5 тыс. лет), Тепекспан (11 тыс. лет). Почти в каждом из этих местонахождениях обнаружены останки нескольких индивидов, часто разного возраста и пола, в большинстве случаев использован " onmouseout="nd();" href="javascript:void(0);">кроманьонцы
были пришельцами, то откуда они пришли? Как они взаимодействовали с аборигенами - Неандертальцы
- часть неандертальцев жила в суровых условиях ледникового периода, что повлекло сильную биологическую и культурную специализацию.");" onmouseout="nd();" href="javascript:void(0);">неандертальцами
?
Прежде всего, стоит сказать о судьбе европейских неандертальцев. Раньше считалось, что они эволюционировали в людей современного типа, одна стадия перешла в другую. Такое мнение возникло еще тогда, когда были известны лишь европейские находки. Сейчас такой сценарий кажется почти невероятным - слишком велики различия в строении и культуре, да и сосуществование неандертальцев и кроманьонцев уже доказано. Возможно, неандертальцы вымерли или были истреблены кроманьонцами? Однако неандертальцы были лучше приспособлены к условиям ледникового периода, особенно учитывая тропическое происхождение кроманьонцев. До этого неандертальцы много тыс. лет жили на этой территории и были как нельзя лучше адаптированы именно к такой окружающей среде. Да и физически они были намного сильнее кроманьонцев. Некоторые ученые считают, что в борьбе за территории кроманьонцам помог неизмеримо более высокий уровень технического оснащения и социальной организации. Кроме того, вовсе не исключено смешение некоторых групп первых Неоантропы
- стадия человеческой эволюции, включающая и современного человека. Соответствуют ситематическому названию Homo sapiens sapiens
. Во времени следуют за палеоантропами, то-есть, согласно стадиальной теории, появились около 40 тысяч лет назад. Отличаются большим мозгом, наличием выступающего подбородка, очень сложным поведением, включающем язык и искусство. Культура неоантропов времени от 40 до 10 тысяч лет назад называется верхний палеолит. По современным данным, люди современного облика жили уже около 100 тысяч лет назад и были носителями культуры мустье.");" onmouseout="nd();" href="javascript:void(0);">неоантропов
и поздних неандертальцев. Об этом свидетельствуют находки скелетов с промежуточными признаками, вероятно - метисов неандертальцев и кроманьонцев. Можно также вспомнить о чертах Мустье
- средний палеолит, техника обработки камня или комплекс культур, возникший около 200 и исчезнувший 35 тысяч дет назад. Типичными орудиями являются остроконечник и скребло. Носителями техники мустье были палеоантропы и часть ранних неоантропов. Обычно с мутье не встречается признаков искусства и костяных орудий, что свидетельствует о низком уровне культуры и абстрактного мышления его создателей.");" onmouseout="nd();" href="javascript:void(0);">мустье
в раннем верхнем палеолите Европы и чертах верхнего палеолита на некоторых мустьерских стоянках. А с культурой шательперрон связаны остатки как поздних неандертальцев, так и ранних кроманьонцев. Вероятно, именно такое смешение генов и культур помогло первым неоантропам быстро приспособиться к совершенно новым для них природным условиям. И не с тех ли пор у европейцев относительно широкая кисть, широкая стопа, массивные, сравнительно с населением других материков, череп и скелет? (см. Хрестом. 4.5).
Сейчас нет однозначного ответа на вопрос о судьбе неандертальцев. Новые исследования еще прольют свет на эту интересную проблему, а пока, предлагаем Вам еще раз рассмотреть таксономическую систему семейства Гоминиды
- семейство приматов, к которому относят современного человека. По наиболее распространенной схеме включают два подсемейства - австралопитецин и гоминин. Некоторые ученые считают, что к гоминидам можно отнести и человекообразных обезьян.");" onmouseout="nd();" href="javascript:void(0);">гоминид
, большинство видов и родов которого, кроме Homo sapiens, являются вымершими (табл. 4.6).
| Название (первое описание) | Синонимы | Регион | Время, млн. л. |
|---|---|---|---|
| Ранние австралопитеки | |||
| Sahelanthropus tchadensis (Brunet et al., 2002) | - | Чад | 6-7 |
| Orrorin tugenensis (Senut et al., 2001) | - | Кения | 5,6-6,2 |
| Ardipithecus ramidus (White et al., 1995) | =Homo antiquus praegens
(Ferguson, 1989) =Australopithecus praegens |
Эфиопия | 3,9-5,5 |
| Australopithecus anamensis (Leakey et al., 1995) | - | Кения | 3,9-4,2 |
| Грацильные австралопитеки | |||
| Australopithecus afarensis (Taieb et al., 1978)(возможно, 2 вида) | =Australopithecus africanus tanzaniensis
(Tobaios, 1980) =Australopithecus aethiopicus (Tobias, 1980) =Homo antiquus (Ferguson, 1984) =Meganthropus africanus (Weinert, 1950) =Praeanthropus africanus (Hennig, 1948; sensu Senuyrek, 1955) =Paranthropus africanus (Weinert, 1950; sensu Olson, 1981) |
Кения, Танзания, Эфиопия | 3,0-3,9(4,4) |
| Kenyanthropus platyops (Leakey et al., 2001) | - | Кения, Эфиопия | 3,2-3,5 |
| Australopithecus bahrelghazali (Brunet et al., 1996) | - | Чад | 3,0-3,5 |
| Australopithecus africanus (Dart, 1925)(возможно, 2 вида) | =Australopithecus prometheus
(Dart, 1948) =Plesianthropus transvaalensis (Broom, 1936) =Homo transvaalensis (Mayr, 1950) |
Южная и Восточная Африка | 2,4-3,5 |
| Australopithecus garhi (Asfaw et al., 1999) | - | Эфиопия | 2,5 |
| Массивные австралопитеки | |||
| Paranthropus aethiopicus (Arambourg et Coppens, 1968) | =Australopithecus walkeri
(Ferguson, 1989) =Paraustralopithecus aethiopicus (Arambourg et Coppens, 1968) |
Кения, Эфиопия | 2,3-2,6 |
| Paranthropus boisei (Leakey, 1959) | =Zijanthropus boisei (Leakey, 1959) | Восточная Африка | 1,1-2,3(2,5) |
| Paranthropus robustus (Broom, 1939)(возможно, 2 вида) | =Paranthropus crassidens (Broom, 1948) | Южная Африка | 0,9-2,5 |
| "Ранние Homo" | |||
| Homo (или Australopithecus) habilis (Leakey et al., 1964) | =Australopithecus habilis
=Homo erectus habilis (Campbell, 1965) ="Prezinjanthropus" =Homo microcranous (Ferguson, 1995) =Telanthropus capensis (Broom, 1947) |
Восточная и Южная Африка | 1,5-2,3-2,6 |
| Homo (или Pithecanthropus) rudolfensis (Alexeev, 1978) | =Kenyanthropus rudolfensis | Восточная Африка | 1,5-2,3(2,5) |
| Род Homo | |||
| Homo (или Pithecanthropus) ergaster (Groves et Mazak, 1975) | =Homo kenyaensis
(Zeitoun) =Homo okotensis (Zeitoun) =Homo georgicus (Gabunia, Vekua, de Lumley et Lordkipanidze, 2002) =Telanthropus capensis (Broom, 1947) |
Африка, Грузия | 1,4-1,7 млн.л. |
| Homo (или Pithecanthropus) erectus (Dubois, 1894)(архантропы) | =Sinanthropus pekinensis
(Black, 1927) =Pithecanthropus leakeyi (Heberer, 1963) =Meganthropus palaeojavanicus (Koenigswald, 1941) =Atlanthropus mauritanicus (Arambourg, 1955) и др. |
Африка, Европа, Азия | 0,4-1,4 млн.л. |
| Homo (или Pithecanthropus) heidelbergensis (Schoetensack, 1908) | =Cyphanthropus rhodesiensis
(Woodward, 1921) =Javanthropus soloensis (Oppenoorth, 1932) и др. |
Африка, Европа, Азия | 130-500 тыс.л. |
| Homo neanderthalensis (King, 1864)(палеоантропы) | =Homo sapiens krapinensis
(Goryanovic-Kramberger, 1902) =Homo mousteriensis (Klaatsch et Hauser, 1909) =Palaeoanthropus palestinensis (Keith et Mc Cown, 1939) и др. |
Африка, Европа, Азия | 35-130(200) тыс.л. |
| Homo sapiens sapiens (Linnaeus, 1758)(неоантропы) | =Neoantropus fossilis
=Homo Africanthropus helmei (Dreyer, 1935) =Homo aurignacensis hauseri (Klaatsch et Hauser, 1912) и др. |
вся планета | 0-40(200)тыс.л. |
4.2. Моноцентризм, дицентризм и полицентризм: сколько у нас предков?
Откуда же взялись люди современного облика? В каких областях Земли происходило сложение современного комплекса биологического строения и поведения - Сапиентация
- процесс возникновения современного вида человека Homo sapiens sapiens
. Сопровождался как биологическими изменениями - увеличением мозга, округлением черепа, уменьшением размеров лица, появлением подбородочного выступа, - так и социокультурными новациями - возникновением искусства, символического поведения, техническим прогрессом, развитием языков.");" onmouseout="nd();" href="javascript:void(0);">сапиентация
?
Изучая древних людей, исследователи давали отличающиеся ответы на эти вопросы. Эти взгляды получили название гипотез Моноцентризм в антропологии
- гипотеза происхождения человека современного вида и его рас в одном географическом центре. Центр этот чаще помещают в Африке. Более умеренным является вариант т.н. широкого моноцентризма, согласно которому центр захватывал также Ближний Восток, Южную и Восточную Европу и часть Средней Азии.");" onmouseout="nd();" href="javascript:void(0);">моноцентризма
, Дицентризм
- гипотеза происхождения современного человечества в двух основных центрах сапиентации - восточном и западном. Оценки времени существования центров и особенностей распространения людей из них несколько различаются у разных авторов.");" onmouseout="nd();" href="javascript:void(0);">дицентризма
и Полицентризм
- гипотеза происхождения современного человека и его рас в нескольких независимых географических центрах. Следует различать строгий полицентризм или полигению, согласно которой разные расы произошли от разных видов ископаемых гоминид, и своременный вариант полицентризма - мультирегиональную гипотезу, согласно которой единство вида поддерживалось генетическими потоками между популяциями. В настоящее время полигения практически не имеет сторонников, тогда как мультирегиональная концепция достаточно популярна.");" onmouseout="nd();" href="javascript:void(0);">полицентризма
, отличие между которыми состоит в разной оценке степени архаичности или прогрессивности географических и хронологических групп древнего населения.
1. Моноцентризм.
Часть антропологов считает, что человек современного облика произошел в достаточно ограниченном регионе планеты. Потом он расселился оттуда по всей Земле, в чем ему помогал высокий уровень интеллекта и значительная биологическая и социальная лабильность. Такой подход к происхождению человека называется моноцентризмом. Очаг возникновения современного человека предполагался в разных областях - Африке, Европе, Центральной Азии, Дальнем Востоке, гипотетической Лемурии и даже в Южной Америке. Гипотеза существования пре-сапиенса в Европе, предшествовавшего неандертальцам, была убедительно подвергнута критике с биологической стороны. Лемурийская и южноамериканская версии явно страдают отсутствием хоть каких-то доказательств. В настоящее время моноцентристы уверенно говорят об Африке (или даже конкретно о Центральной Африке) как о единственном возможном месте возникновения человека. В несколько расширенном варианте речь идет об афро-европейской прародине, включающей также Южную Европу.
Альтернативой может служить так называемая Моноцентризм в антропологии
- гипотеза происхождения человека современного вида и его рас в одном географическом центре. Центр этот чаще помещают в Африке. Более умеренным является вариант т.н. широкого моноцентризма, согласно которому центр захватывал также Ближний Восток, Южную и Восточную Европу и часть Средней Азии.");" onmouseout="nd();" href="javascript:void(0);">гипотеза широкого моноцентризма
, выдвинутая советским антропологом Я.Я. Рогинским . Она предполагает значительно большую территорию, охваченную процессом сапиентации - все Средизмноморье, включая Южную Европу и Северо-Восточную Африку, Ближний Восток и Кавказ (). Процесс сапиентации был, конечно же, не одномоментным и потому завершился на значительно большей территории, нежели начался. Расселяясь из единого центра, Неоантропы
- стадия человеческой эволюции, включающая и современного человека. Соответствуют ситематическому названию Homo sapiens sapiens
. Во времени следуют за палеоантропами, то-есть, согласно стадиальной теории, появились около 40 тысяч лет назад. Отличаются большим мозгом, наличием выступающего подбородка, очень сложным поведением, включающем язык и искусство. Культура неоантропов времени от 40 до 10 тысяч лет назад называется верхний палеолит. По современным данным, люди современного облика жили уже около 100 тысяч лет назад и были носителями культуры мустье.");" onmouseout="nd();" href="javascript:void(0);">неоантропы
смешивались с аборигенными популяциями Палеоантропы
- стадия гоминидной эволюции, следующая за архантропами и предшествующая неоантропам. От архантропов отличаются большим мозгом, от неоантропов - скошенным подбородком, вытянутой формой черепа и значительной массивностью. Европейские и некоторые азиатские палеоантропы также называются неандертальцами Homo sapiens neanderthalensis
или Homo neanderthalensis
. Абычно ассоциированы с ашельской и мустьерской культурой. Искусство у палеонтропов отсутствовало или находилось в зачаточном состоянии.");" onmouseout="nd();" href="javascript:void(0);">палеоантропов
, живших в отдаленных регионах планеты. Этим Я.Я. Рогинский объяснял известное противоречие - наличие одновременно сходных признаков у современных людей очень отдаленных территорий и сходство в отдельных признаках современных рас с ископаемыми гомининами тех же областей.
2. Полицентризм. Последняя особенность - географическое распределение специфических признаков - привела к появлению Полицентризм - гипотеза происхождения современного человека и его рас в нескольких независимых географических центрах. Следует различать строгий полицентризм или полигению, согласно которой разные расы произошли от разных видов ископаемых гоминид, и своременный вариант полицентризма - мультирегиональную гипотезу, согласно которой единство вида поддерживалось генетическими потоками между популяциями. В настоящее время полигения практически не имеет сторонников, тогда как мультирегиональная концепция достаточно популярна.");" onmouseout="nd();" href="javascript:void(0);">полицентризма . Опираясь на сходство современных людей и ископаемых гоминин тех же областей (монголоидов и синантропов - в особой лопатовидной форме резцов, неандертальцев и европейцев - в силовом типе кисти, африканцев и черепа из Брокен Хилл в Замбии - в больших размерах челюстей и т.п.), некоторые ученые предположили независимое происхождение разных человеческих рас. Основателем концепции полицентризма является немецкий антрополог Ф. Вейденрейх .
- Он выделил четыре центра формирования современных рас (1941):
- Европеоиды - представители европеоидной расы (в разных классификациях ее называют кавказоидной, или евразийской расой), распространенной в Европе, Северной Африке (до Сахары), Ближнем Востоке, Средней и Центральной Азии, Северной Индии. Характерные признаки населения: цвет кожи, волос и глаз варьирует от очень светлых оттенков у северных групп до очень темных у южных и восточных, волосы прямые или волнистые, мягкие, лицо ортогнатное, в горизонтальной плоскости (при взгляде сверху) заметно выступает вперед, надбровные дуги часто большие, орбиты низкие, угловатые, разрез глаз всегда широкий, но глазная щель может быть небольшой, эпикантуса нет, нос обычно крупный, резко выступает, ноздри ориентированы сагиттально, переносье высокое, рот неширокий, толщина губ небольшая или средняя, рост бороды и усов сильный, телосложение среднее, кисть и стопа широкие. На краях ареала европеоидная раса образует плавные переходы к уральской, южносибирской, монголоидной, негроидной, эфиопской и дравидийской расам.");" onmouseout="nd();" href="javascript:void(0);">европеоиды возникли в Южной и Центральной Европе;
- Негроид - представитель популяции негроидной расы.");" onmouseout="nd();" href="javascript:void(0);">негроиды - в Африке;
- Монголоидная (или азиатская) раса (монголоиды) - распространена в основном в Азии. Иногда объединяется с американоидной расой в большую азиатско-американскую расу. Характерные черты собственно монголоидной расы (без американоидной): уплощенное лицо с выдающимися скулами, чаще высокое, высокие орбиты, цвет волос и глаз почти всегда черный, цвет кожи от светлого у северных групп до смугловатого у южных, но никогда не бывает очень темным, ортогнатизм и мезогнатизм, волосы обычно прямые или слабоволнистые, рост бороды, усов и волос на теле очень слабый, разрез глаз узкий, часто встречается эпикантус, степень выступания носа варьирует, но чаще небольшая, переносье обычно вогнутое, толщина губ от малой до средней, ширина рта небольшая, рост в мировом масштабе невысокий, пропорции коренастые, ноги относительно укорочены. На границах ареала имеет плавные переходы в европеоидную, дравидийскую, веддоидную, меланезийскую, полинезийскую, уральскую и южносибирскую расы.");" onmouseout="nd();" href="javascript:void(0);">монголоиды - в Восточной Азии;
- Австралоидная (или австралийская) раса (австралоиды) - коренное население Австралии. Характерные черты: большая массивность долихокранного черепа при небольшой массивности остального скелета, сильные надбровные дуги, крупные прогнатные челюсти, большие зубы, короткая шея, очень темная пигментация кожи, волос и глаз (иногда глыбки пигмента даже выходят за пределы радужины; однако у некоторых групп Центральной Австралии встречается светлая окраска волос), очень широкий нос со сравнительно высоким переносьем, глубоко посаженные глаза, волосы волнистые (на юго-западе в Виктории есть курчавоволосые группы, возможно, это следствие смешения с тасманийцами, которые во множестве переселялись в порт Филиппа в ранний период европейской колонизации), рост бороды и усов сильный, отмечен особый тип направления волос на теле, рост выше среднего и высокий, телосложение очень вытянутое. Иногда выделяют в самостоятельную большую расу, иногда объединяют с негроидами в австрало-негроидную или экваториальную расу, по признакам строения зубов относят к восточному надрасовому стволу. Наиболее близки к австралоидной расе веддоидная и меланезийская расы, с которыми иногда объединяется в австрало-веддоидную расу.");" onmouseout="nd();" href="javascript:void(0);">австралоиды - на Больших Зондских островах.
Развил эту гипотезу К. Кун, разделивший африканский очаг расообразования на два независимых - северный и южный, в которых возникли соответственно капоиды и негроиды, переселившиеся затем в противоположные части континента. По К. Куну, каждый из пяти очагов был населен собственным подвидом архантропов. Современные же расы он делил на более и менее развитые ().
Несколько более умеренный вариант полицентризма с выделением 3-4 центров Сапиентация
- процесс возникновения современного вида человека Homo sapiens sapiens
. Сопровождался как биологическими изменениями - увеличением мозга, округлением черепа, уменьшением размеров лица, появлением подбородочного выступа, - так и социокультурными новациями - возникновением искусства, символического поведения, техническим прогрессом, развитием языков.");" onmouseout="nd();" href="javascript:void(0);">сапиентации
, близких друг к другу, предложил венгерский антрополог А. Тома. В отличие от К. Куна, А. Тома считал уровень современных рас одинаковым ().
Полицентрические концепции защищали также отечественные антропологи и археологи - Г.Ф. Дебец , В.П. Алексеев , Ю.И. Семенов, А.А. Формозов и ряд других. Однако они исходили из принципиально других соображений, нежели западные ученые. Непрерывность развития палеоантропов в неоантропов на разных территориях обосновывалась влиянием труда и законами развития общества, едиными для всех человеческих коллективов. Миграция же неоантропов из единого центра возникновения слишком напоминала мысль о завоевании жизненного пространства "высшей расой".
В современной зарубежной науке гипотеза полицентризма переросла в Мультирегиональная гипотеза
- вариант полицентризма, предполагающий сущестсвование нескольких центров сапиентации, между которыми происходил обмен миграционными потоками, вследствие чего сохранялось единство вида.");" onmouseout="nd();" href="javascript:void(0);">мультирегиональную гипотезу
. Ее сторонники стараются особенно подчеркнуть два основных момента: преемственность биологических признаков от архантропов до современного человека в Африке, Европе и Юго-Восточной Азии и Австралии, с одной стороны, и неравнозначность терминов "Полицентризм
- гипотеза происхождения современного человека и его рас в нескольких независимых географических центрах. Следует различать строгий полицентризм или полигению, согласно которой разные расы произошли от разных видов ископаемых гоминид, и своременный вариант полицентризма - мультирегиональную гипотезу, согласно которой единство вида поддерживалось генетическими потоками между популяциями. В настоящее время полигения практически не имеет сторонников, тогда как мультирегиональная концепция достаточно популярна.");" onmouseout="nd();" href="javascript:void(0);">полицентризм
" и "полигенность" - с другой. По мнению современных полицентристов, единство человеческого вида всегда поддерживалась путем многочисленных миграций - генных и культурных потоков между регионами. Так сохранялось и единство, и разнообразие человечества.
3. Дицентризм.
Вариантом полицентризма является Дицентризм
- гипотеза происхождения современного человечества в двух основных центрах сапиентации - восточном и западном. Оценки времени существования центров и особенностей распространения людей из них несколько различаются у разных авторов.");" onmouseout="nd();" href="javascript:void(0);">дицентризм
. Наиболее стройный вариант дицентризма предложен отечественным антропологом А.А. Зубовым (; ;). Проанализировав мировое распределение одонтологических признаков, он предположил, что человечество на стадии Архантропы
- группа или стадия ископаемых гоминид, существовавшая после " homo="">Homo erectus. Архантропы жили примерно от 1,6 до 400 тысяч лет назад.");" onmouseout=nd(); href="javascript:void(0);">архантропов
делилось на два главных расовых ствола - восточный (монголоиды и австралоиды) и западный (европеоиды и негроиды). Особенность такого подхода заключается в том, что признаки зубов имеют высокую генетическую детерминацию, напрямую не зависят от условий окружающей среды и потому не подвержены прямому действию Естественный отбор
- термин, введенный Ч. Дарвином (1858 г.), для обозначения явления преимущественного выживания и оставления потомства наиболее приспособленными к условиям обитания особями данного вида, и гибели менее приспособленной части популяции в результате борьбы за существование. Является основным движущим фактором эволюции: более выгодные для популяции признаки, способствующие выживанию особей, передаются в следующее поколение. Независимо от Ч. Дарвина к этой идее пришел А. Уоллес также в 1858 г.");" onmouseout="nd();" href="javascript:void(0);">естественного отбора
. Они возникают достаточно случайно. Следовательно, они могут быть хорошим индикатором древности группы и степени ее изолированности от других популяций. По этим признакам А.А. Зубов выделяет два очага сапиентации, которые оказались изолированными в глубокой древности. В последующем тропические популяции обоих стволов вследствие одинаковых условий приобрели значительное физиономическое сходство и ныне выделяются в единую экваториальную расу.
В.П. Алексеев предположил несколько иной вариант развития событий: позднюю миграцию части экваториалов западного ствола на восток, в результате чего сложилась австралоидная раса (). Таким образом, два центра происхождения современного вида человека у В.П. Алексеева сохраняются, но экваториальная раса оказывается возникшей из одного центра - западного (см. Хрестом. 4.6).
Аргументы моноцентристов и полицентристов многочисленны и подтверждаются фактическим материалом. Однако они никак не могут перевесить друг друга, и потому пока нельзя твердо ответить на вопрос - где же возник современный человек? Вы можете выбрать себе наиболее понравившеюся концепцию - все они на настоящий момент являются вполне убедительными и равнозначными.
4.3. Резюме
- Итак, подведем некоторые итоги:
- Согласно палеонтологическим данным, линия предшественников, ведущая к человеку, отделилась от линии современных человекообразных обезьян около 4-8 миллионов лет назад.
- Существует огромное количество находок ископаемых приматов, выстраивающихся в последовательный Эволюционный ряд - последовательный ряд форм организмов (ископаемых и(или) современных), направление изменения которых выстраивается в логичную прямую последовательность. При помощи эволюционных рядов устанавливается филогенетическое родство организмов и их систематика.");" onmouseout="nd();" href="javascript:void(0);">эволюционный ряд форм - от обезьян до человека.
- Ответ на вопрос - когда и где возник человек? - зависит от того, кого мы считаем человеком, а кого - нет.
- В науке есть множество взаимоисключающих мнений по поводу причин, времени и места возникновения современного человека. Нерешенных вопросов становится тем больше, чем больше мы даем ответов на предыдущие вопросы.
- Существовало огромное количество предшественников современного человека, не все из которых были нашими предками. Их можно разделять на Стадия - крупный этап развития чего-либо (как правило, подразумевается, что в течении этого этапа объект изменяется не сильно).");" onmouseout="nd();" href="javascript:void(0);">стадии или выделять группы согласно биологической Систематика - раздел общей биологии, призванный создать единую стройную систему животного мира на основе выделения системы биологических таксонов и соответствующих названий, выстроенных по определенным правилам (номенклатура). В идеале систематика должна отражать филогению, но это далеко не просто в связи с чем, является одним из наиболее запутанных разделов науки.");" onmouseout="nd();" href="javascript:void(0);">систематике (табл. 4.6).
- Вместе с биологической эволюцией шла и Эволюция социальная - термин используемый для описания направленных (как правило, прогрессивных) исторических изменений социальной организации человека. В неявном виде говоря так предполагается наличие общих закономерностей социальных изменений (их направленность, причинно-следственная обусловленность).");" onmouseout="nd();" href="javascript:void(0);">эволюция социальная , реконструируемая, в частности, методами Археология (от греч. archaios древний и logos наука) - историческая дисциплина, исследующая прошлое человечества по памятникам материальной культуры, с использованием специальных приемов и методов (основной из них - проведение археологических раскопок).");" onmouseout="nd();" href="javascript:void(0);">археологии .
- Согласно наиболее аргументированной точке зрения, человек современного облика и поведения возник в Африке или на Ближнем Востоке от 100 до 40 тыс. лет назад.
- Со времени около 35 тыс. лет на Земле существует только современный вид человека, распространившийся по всей планете.
Словарь терминов
- Гоминиды - семейство приматов, к которому относят современного человека. По наиболее распространенной схеме включают два подсемейства - австралопитецин и гоминин. Некоторые ученые считают, что к гоминидам можно отнести и человекообразных обезьян.");" onmouseout="nd();" href="javascript:void(0);">Гоминиды
- Дицентризм - гипотеза происхождения современного человечества в двух основных центрах сапиентации - восточном и западном. Оценки времени существования центров и особенностей распространения людей из них несколько различаются у разных авторов.");" onmouseout="nd();" href="javascript:void(0);">Дицентризм
- Моноцентризм в антропологии - гипотеза происхождения человека современного вида и его рас в одном географическом центре. Центр этот чаще помещают в Африке. Более умеренным является вариант т.н. широкого моноцентризма, согласно которому центр захватывал также Ближний Восток, Южную и Восточную Европу и часть Средней Азии.");" onmouseout="nd();" href="javascript:void(0);">Моноцентризм
- Мультирегиональная гипотеза - вариант полицентризма, предполагающий сущестсвование нескольких центров сапиентации, между которыми происходил обмен миграционными потоками, вследствие чего сохранялось единство вида.");" onmouseout="nd();" href="javascript:void(0);">Мультирегиональная гипотеза
- Полицентризм - гипотеза происхождения современного человека и его рас в нескольких независимых географических центрах. Следует различать строгий полицентризм или полигению, согласно которой разные расы произошли от разных видов ископаемых гоминид, и своременный вариант полицентризма - мультирегиональную гипотезу, согласно которой единство вида поддерживалось генетическими потоками между популяциями. В настоящее время полигения практически не имеет сторонников, тогда как мультирегиональная концепция достаточно популярна.");" onmouseout="nd();" href="javascript:void(0);">Полицентризм
- Сапиентация - процесс возникновения современного вида человека Homo sapiens sapiens . Сопровождался как биологическими изменениями - увеличением мозга, округлением черепа, уменьшением размеров лица, появлением подбородочного выступа, - так и социокультурными новациями - возникновением искусства, символического поведения, техническим прогрессом, развитием языков.");" onmouseout="nd();" href="javascript:void(0);">Сапиентация
- Сетевидная эволюция - теоретический вариант хода эволюционного процесса, при котором новые признаки возникают в разных популяциях, объединяясь в комплексы через смешение этих популяций. Сторонники сетевидной эволюции придают большое значение миграции, как эволюционному фактору.");" onmouseout="nd();" href="javascript:void(0);">Сетевидная эволюция
- Стадиальная теория антропогенеза - теория антропогенеза, утверждающая, что в ходе эволюции человека последовательно сменялись несколько стадий предков человека: австралопитеки (протантропы) - архантропы - палеоантропы - неоантропы. Одновременно менялся как биологический облик, так и поведение: олдувайская культура сменилась ашельской, потом мустьерской, затем верхним палеолитом, который перешел в мезолит, неолит, бронзовый и, наконец, железный век. Сейчас стадиальная теория имеет скорее историческое значение, поскольку ясно, что в антропогенезе сосуществование примитивных и прогрессивных групп было распространено очень широко.");" onmouseout="nd();" href="javascript:void(0);">Стадиальная теория антропогенеза
Вопросы для самопроверки
- Приведите отличия и сходство приматов и человека (биологические и небиологические). Какая "грань" отделяет человека от обезьяны?
- Что такое "гоминидная триада", какие другие биологические и небиологические признаки можно считать уникальными для гоминид?
- Можно ли говорить о "моменте возникновения" современного человека, учитывая при этом данные палеоантропологии?
- В какой последовательности возникали биологические и поведенческие особенности современного человека?
- Кого считают предками современного человека с позиций эволюционной биологии? (При ответе постарайтесь рассмотреть разные гипотезы)
- В чем заключаются концепции моно- и полицентризма, почему в данный момент они считаются равноправными?
- Что такое мультирегиональная гипотеза происхождения человека?
- Что такое "прародина" человека?
- В чем заключается стадиальная гипотеза происхождения человека? Какие другие концепции выдвигает современная антропология?
- Как можно исследовать поведение предшественников человека? Чем поведение современного человека отличается от поведения ископаемых людей разных эпох?
Темы курсовых работ и рефератов
- Корни возникновения человека - недостающие звенья.
- Австралопитеки - обезьянолюди или человекообезьяны?
- Критерии культуры и ее начало.
- Архантропы - антропология и культура.
- К проблеме близости культуры и биологии палеоантропов и людей современного вида.
- Люди верхнего палеолита - возникновение современной психики.
- Центры сапиентации - число и время возникновения.
- Развитие культуры и психики древнего человека.
- К вопросу о динамике развития психики и поведения предков человека по данным археологии и антропологии.
Список литературы
Обязательная
- Алексеев В.П. Становление человечества. М. 1984.
- Ископаемые гоминиды и происхождение человека / Под ред. В.В. Бунака // Труды Института этнографии АН СССР. Новая серия. Т. 92. М., 1966.
- Рогинский Я.Я., Левин М.Г. Антропология. М., 1963.
- Хрисанфова Е.Н., Перевозчиков И.В. Антропология: Учебник. 2-е изд. М., 1999.
- Алексеев В.П. Человек. Эволюция и таксономия (некоторые теоретические вопросы). М., 1985.
- Алексеев В.П. Палеоантропология земного шара и формирование человеческих рас. Палеолит. М., 1978.
- Бунак В.В. Род Homo, его возникновение и последующая эволюция. М., 1980.
- Бутовская М.Л., Файнберг Л.А. У истоков человеческого общества (Поведенческие аспекты эволюции человека). М., 1993.
- Джохансон Д., Иди М. Люси. Истоки развития человеческого рода. М., 1984.
- Зубов А.А. Иерархия факторов антропогенеза // Вестник антропологии. Вып. 2, 1996.
- Зубов А.А. Магистрализация и демагистрализация в ходе эволюционного процесса // Вопросы антропологии. Вып. 75, 1985.
- Зубов А.А. Эволюция рода Homo от архантропа до современного человека // Итоги науки и техники. Серия "Антропология". Т. 2. Становление и эволюция человека. М., 1987.
- Кочеткова В.И. Палеоневрология. М., 1973.
- Оуэн Лавджой К. Эволюция выпрямленного способа передвижения у человека // В мире науки. Scientific American. № 1., 1998.
- Семенов Ю.И. На заре человеческой истории. М., 1989.
- Фоули Р. Еще один неповторимый вид. М. 1990.
- Хрисанфова Е.Н. Древнейшие этапы гоминизации // Итоги науки и техники. Серия "Антропология". Т. 2. Становление и эволюция человека. М. 1987.
- Энгельс Ф. Роль труда в процессе превращения обезьяны в человека. М. 1952.
- Coon C.S. The origin of races. New York. 1962.
- Thoma A. Le Peuplement Antйneandertalien de l"Europe. Paris. 1976.
- Weidenreich F. Six lectures on Sinanthropus and related problems // Bulletin of the Geological Society of China. Vol. 19. 1939.
- Weidenreich F. The skull of Sinanthropus pekinensis: a comparative study of a primitive hominid skull // Palaeontologica Sinica. New Series. D. No. 10. 1943.
АНТРОПОГЕНЕЗ
Антропология………………………………………………………...3
Классификация приматов…………………………………………5
Полуобезьяны и обезьяны………………………………………..7
Происхождение человека…………………………………………14
Ископаемые предки человека……………………………………17
Австралопитеки………………………………………………………21
Человек умелый……………………………………………………...23
Архантропы……………………………………………………………25
Неандертальцы……………………………………………………….34
Неоантропы……………………………………………………………41
Доказательства животного происхождения человека……..57
Отличие человека от животных………………………………….61
Механизм антропогенеза……………………….………………….65
Расы …………………………………………………………………….67
Механизм расогенеза……………………………………………….78
Современные первобытные люди……………………………...80
Контрольный тест……………………………………………………83

Антропология
Термин антропология ввел еще Аристотель, применительно к науке о человеке. Антропология – это фактически комплекс наук о человеке , как биологических, так и общественных.
Новое направление эта наука получила благодаря Ч. Дарвину, который доказал животное происхождение человека (от древнейших человекообразных обезьян). В отличии от животных человек является «общественным существом» и основным условием развития его является труд и общество.
Антропология использует методы сопредельных наук, изучает как мертвые останки древних людей, так и живых людей разных этнических, социальных, возрастных и половых групп.
Методы антропологии так же широко используются в жизни людей, так антропометрия (количественное определение различных особенностей тела) используется для оценки физического развития различных групп учащихся, призывников, а соматоскопия (описание признаков) используется в криминалистике для идентификации личности.

Комплекс современных наук, изучающих человека

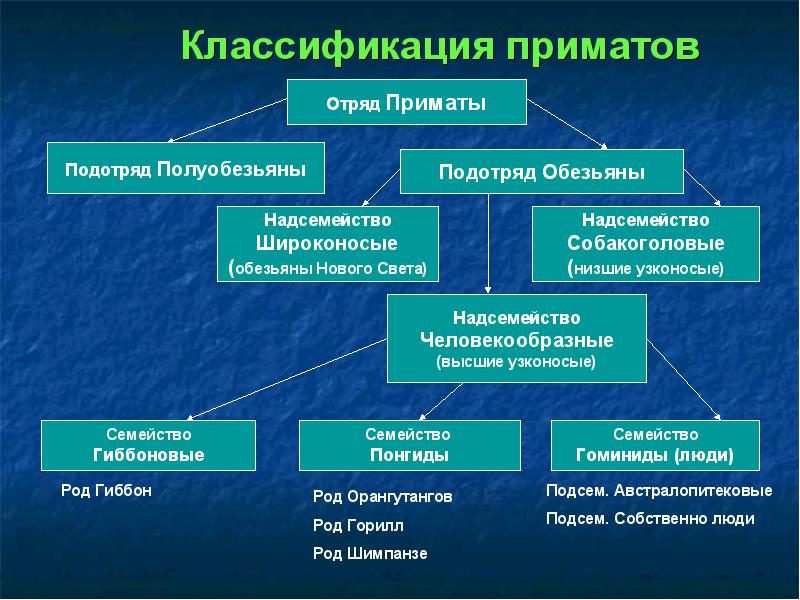
Подсемейство Собственно люди

Лемуры и галаго (полуобезьяны)

ОБЕЗЬЯНЫ СТАРОГО СВЕТА

ОБЕЗЬЯНЫ НОВОГО СВЕТА

ШИМПАНЗЕ
Шимпанзе – наши ближайшие родст-венники, т.е. у нас с ними общие предки. Они отличаются редким для зверей умением пользоваться орудия-ми труда, сообразительностью и способностью к творческой деятельности.
Вытянутое лицо лишено волосяного покрова, светлое.
Обитают в тропических лесах Западной и Центр. Африки. Живут крупными сообществами (до 100 и более особей) со строгой иерархией.
Питаются фруктами, семенами, сочными листьями, цветками , личинками насекомых и охотятся (груп-пой) на детенышей свиней, антилоп и других обезьян..
Размножаются круглый год. Половая зрелость наступает к 7 годам, но спариваться начинают с 13 лет. Детеныши живут с матерью до 7 лет.

ГОРИЛЛА
Гориллы – крупнейшие из всех приматов, несмотря на грозный вид, добродушные вегетарианцы.
Гориллы, как и человек обладают хорошим бинокулярным зрением и глазомером, но из-за большого веса (до 140 кг) взрослые животные живут на земле, а детеныши проводят много времени на деревьях.
Обитают в Центральной Африке. В семействе только один вид – Gorilla gorilla и 3 подвида: Западная береговая, Восточная равнинная и Горная горилла.
Питаются сочными листьями, стеб-лями, корневищами, фруктами, но из-за низкой питательности рациона, они потребляют огромные массы корма, кочуя по участку площадью до 30 км².
Живут группами до 15 особей. Половая зрелость у самок наступает к 10 годам, а у самцов к 15-20 годам.

ОРАНГУТАН
В тропических лесах Азии живет круп-ный представитель приматов – пугли-вый, обособленно живущий орангутан. Две популяции одного вида – Pongo pygmaeus, обитающие на Борнео и Суматре сильно отличаются друг от друга. Суматранский орангутан менее массивен, бледно окрашен и имеет вытянутое лицо.
С помощью длинных рук и ног он очень хорошо передвигается по кронам деревьев. Питаются фруктами, орехами, сочными побегами , иногда листьями, насекомыми, птичьими яйцами и мелкими млекопитающими.
Детеныши сосут грудь до трех лет и остаются с матерью пока та не родит другого малыша. Половой зрелости они достигают к 10-15 годам
Вес взрослых самцов – 70-90 кг, размах рук до 3 м.

ГИББОНЫ
Гиббоны с помощью своих длин-ных рук передвигаются по вет-вям с потрясающей скоростью («летающие обезьяны»).
Питаются фруктами, насекомы-ми, пауками, реже листьями.
Размножаются каждые 3 года. Половозрелость наступает в 6 лет.
Главная особенность гиббонов – ежедневное «пение дуэтом».

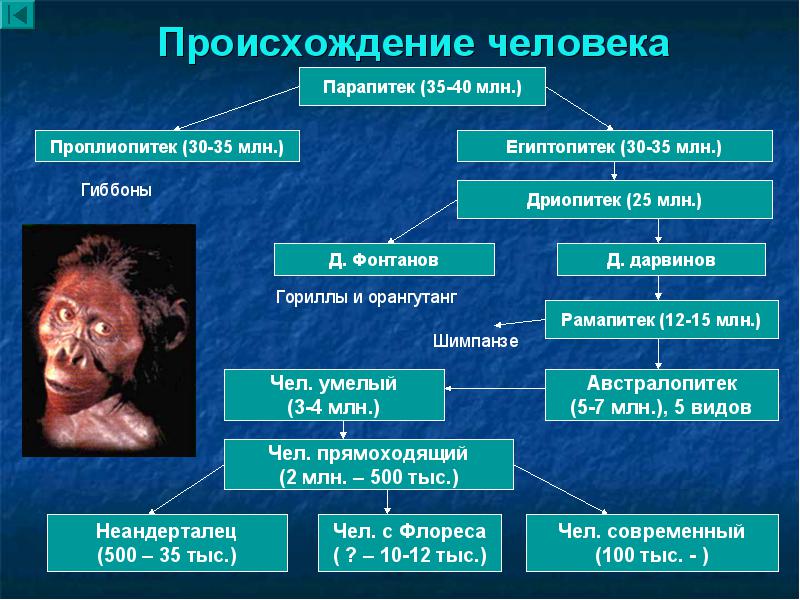
Эволюционное древо гоминидов – надсемейства приматов, включающих 3 семейства: гиббоновых, или малых человекообразных обезьян; понгид, или крупных человекообразных обезьян, и людей.
Проконсул, живший 18 млн. лет назад, является последним общим предком всех понгид и человека, т.к. имеет некоторые черты сходства.

Проконсул – ископаемая обезьяна
Находки, сделанные в 1948, 1951 и 1981 гг. позволили достаточно точно реконструировать этого древнейшего примата. Он имел мозг объемом 160-180 см² и в строении черепа и конечностей напоминал современных понгид.
Жил проконсул около 18 млн. лет назад.
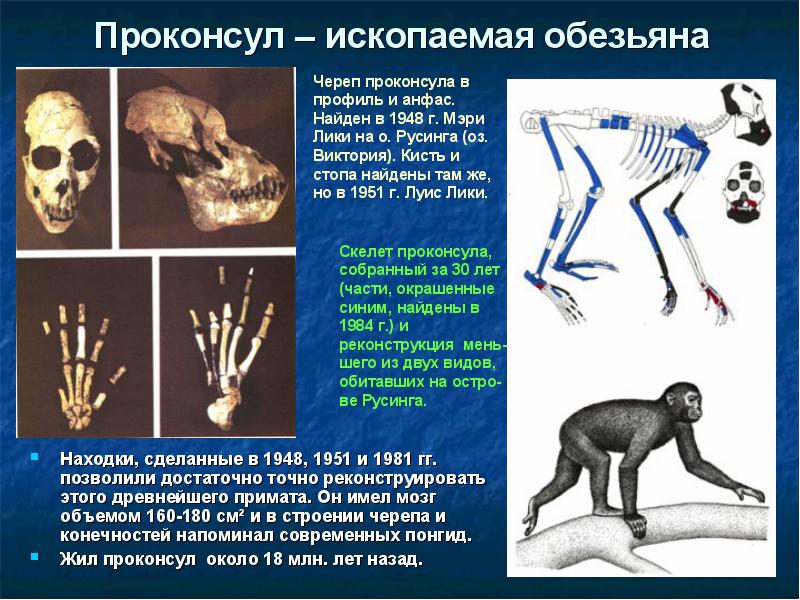
Ископаемые предки человека
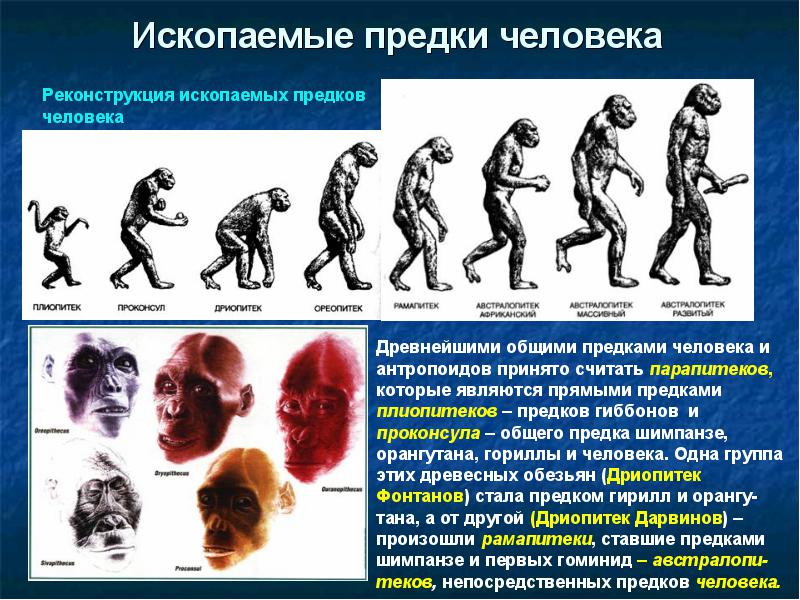
Дриопитек
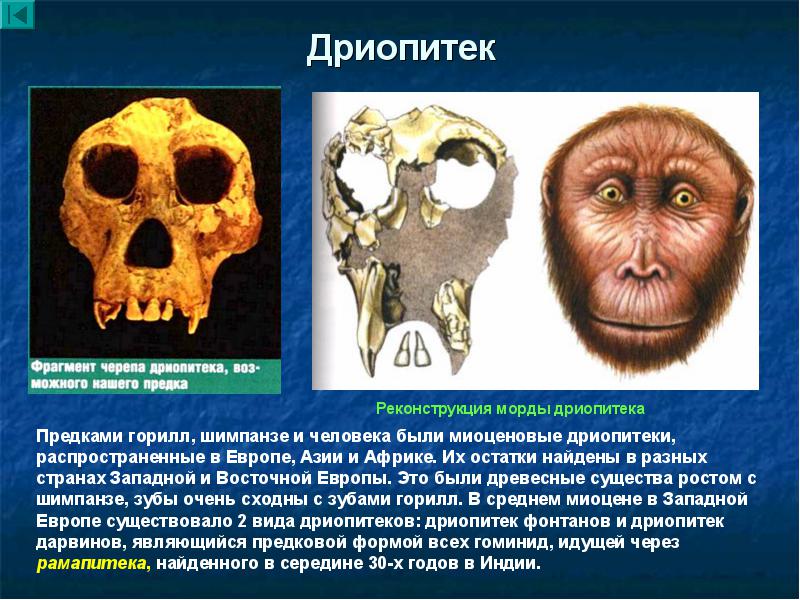
Родословное древо человека

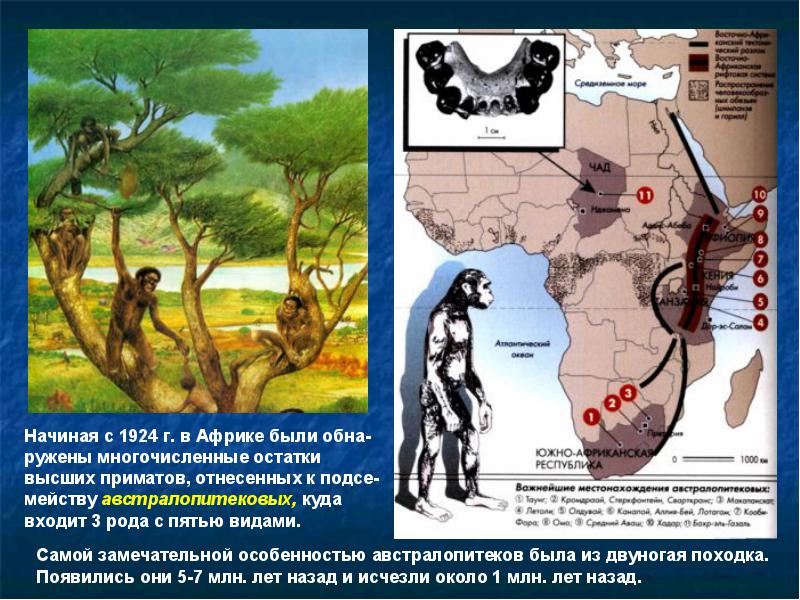
Австралопитеки
В 2002 г. в пустыне Джураб (Чад) найден череп («Туман» - надежда на жизнь) чело-векообразного существа, возраст которого 6-7 млн. лет. Он сочетает в себе черты обезьяны (затылок) и человека (лицо). Это открытие века , т.к. найдена самая ранняя форма гоминид

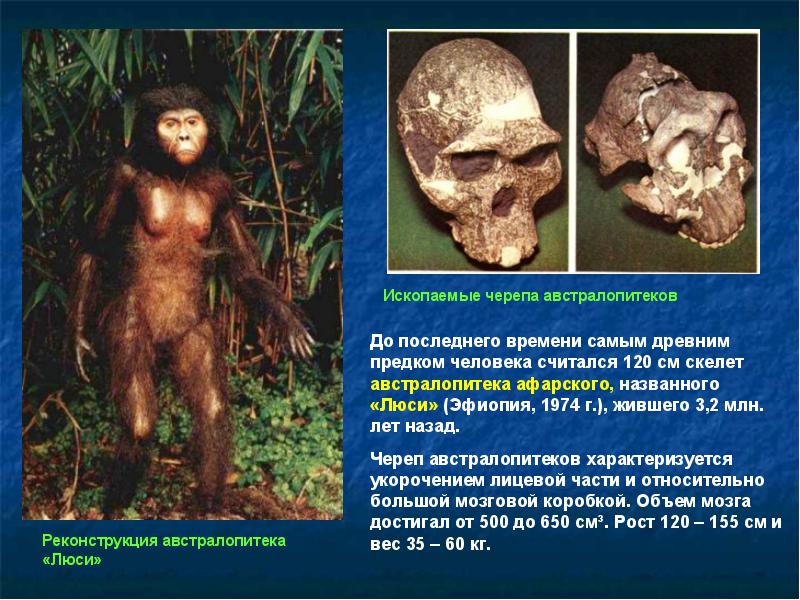
Человек умелый (H. habilis)
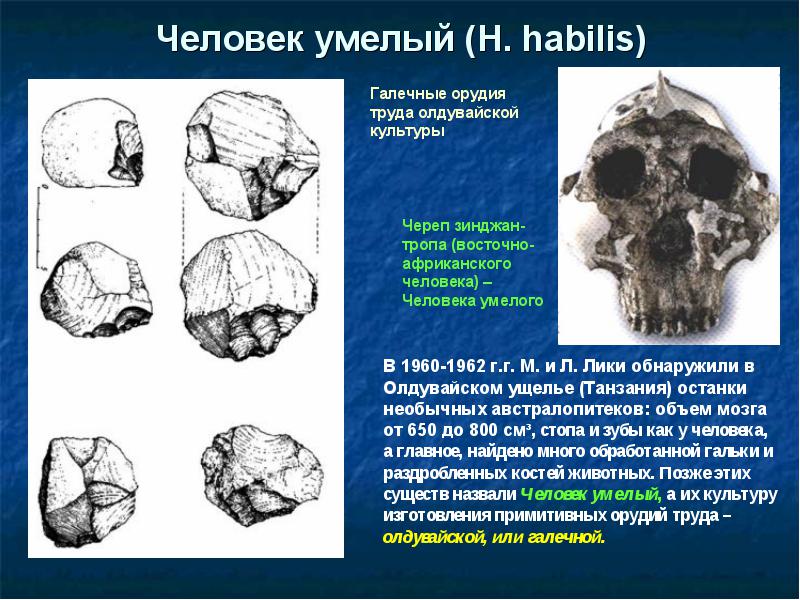
Гипотезы происхождения человека

Архантропы (H. erectus)




Орудия труда архантропов



Вероятный путь расселения человека прямоходящего. 1,5-1,8 млн. лет назад он из Африки переселился на Кавказ, а затем отдельные группы мигрировали в Европу и Ю–В Азию.
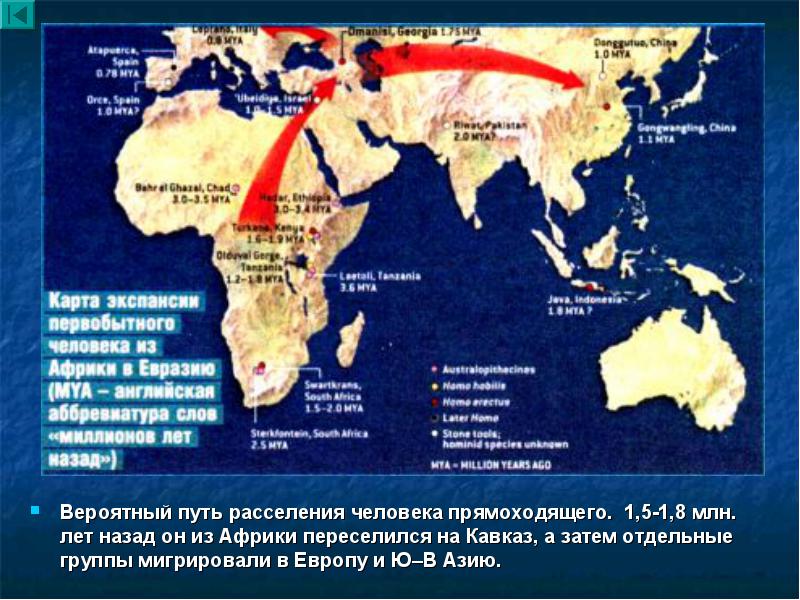
Палеоантропы, или неандертальцы

Неандерталец




Генетические исследования ДНК в найденных костных останках неандертальцев, живших 30 тыс. лет назад показали, что они не являются предками современного человека, а были тупиковой ветвью в развитии гоминид.
Ранние палеоантропы выглядят очень «современно», а поздние более звероподобно, как архантропы.

Как считают английские исследователи генетически рыжий цвет волос, видимо, был унаследован человеком от неандертальцев (ген старше, чем человек современный) при смешанных браках этих двух подвидов примерно 28-35 тыс. лет назад.
В 2004 г. был обнаружены останки 6-7- летнего мальчика, жившего 30 тыс. лет назад, ДНК которого содержит гены неандертальца и человека современного, что подтвер-ждает гипотезу о частичном растворении неандертальцев среди неоантропов.

Неоантропы

небольшой надбровный валик
Неоантропы были высокого роста (170-180 см) с крупными грубыми чертами (сходны с жителями Кавказа и Балкан). Объем мозга – 1200 – 1400 см³. Череп имел высокий лоб, небольшой надбровный валик , легкую нижнюю челюсть с четко выраженным подбородочным выступом, что свидетельствует о развитой членораздельной речи.
Жили они в пещерах или строили жилища. Украшали себя ожерельями, браслетами, кольцами. Характерной особенностью стала наскальная живопись (Капова пещера на Ю. Урале). У них биологические факторы в эволюции утратили свое значение и эволюция приобрела социальный характер.






Неоантропы жили во времена последнего ледникового периода. Как и неандертальцы они охотились на крупных животных, устраивая для них всевозможные ловушки. Мясо одного мамонта могло прокормить одно человеческое племя в течение 2-3 мес., поэтому стоянки устраивались чаще там, где убивали крупное животное (в условиях вечной мерзлоты мясо не портилось, но требовало охраны от диких хищников)

Происхождение человека современного – самая загадочная «глава» антропологии.


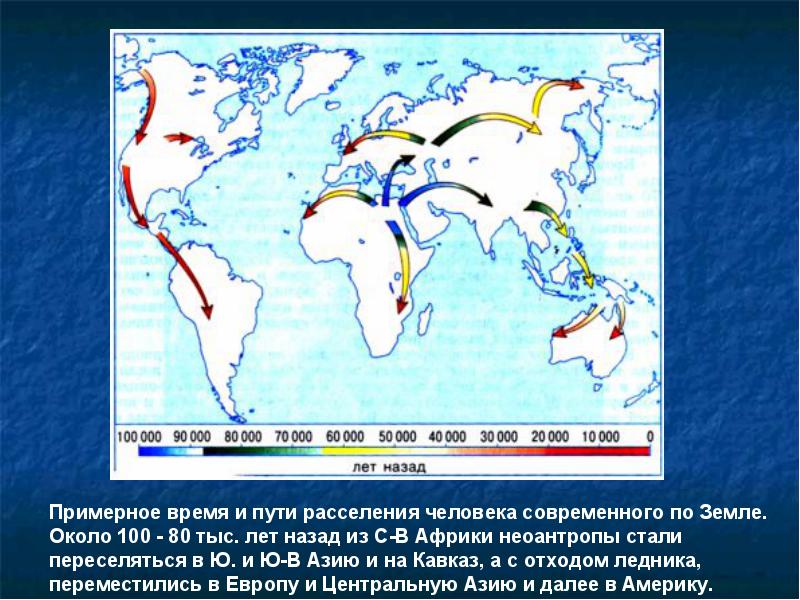
Человек современный был широко распространен в Евразии и Африке и три его крупные географические популяции сформировались в три основные расы: европеоидную (кроманьонцы), монголоидную (человек Верхней пещеры) и негроидную (Боскопский человек).
Прямыми предками кроманьонцев ученые считают басков – древнюю народность Испании, у которых даже язык не имеет родственных корней ни с одним языком мира.
Главным фактором антропо-генеза считают естественный отбор на лучшую орудийную деятельность. В этом жестком отборе победил человек современный.


Ископаемый Человек с о. Флорес

Загадочные находки
На Востоке о. Ява в 1974 г. было обнаружено 2 мумии высохших человечков мужского пола, рост которых не превышал 25 см. У одного была небольшая седая борода. По предварительным данным умерли они около 3 – 4 тыс. лет назад.
Видимо, существующие у разных народов легенды о «гномах» имеют под собой реальную почву.

Доказательства животного происхождения человека
Человек имеет все основные признаки млекопитающих: 1 – левосторонняя дуга аорты; 2 – постоянная температура тела; 3 – эритроциты без ядер ; 4 – две смены зубов, которые разделены на 4 группы 5 – 7 шейных позвонков (нет ни одной кости, какой – бы не было у млекопитающих); 6 – 6-месячный плод покрыт шерстью;

Доказательства родства человека и понгид
Диплоидное число хромосом у шимпанзе – 48, а у человека – 46, но хромосома 2 (см. рис.) образована слиянием двух хромосом шимпанзе.
Дифференциальная окраска хромосом (поперечная полосатость), строго специфичная для каждого вида, удивительно близка у человека и шимпанзе.
Группы крови человека и понгид очень близки, а система АВО у них общая. Кровь бонобо вообще сходна с человеческой.
Одинаковая зубная формула (32 зуба).
Генетическое родство.
Сходная аминокислотная последова-тельность в белках (около 1% замен), а многие белки (гормоны) взаимозаме-няемы.
Молекулярно – генетическая близость человека и понгид свидетельствует, что они должны быть объединены в одно семейство людей (Homo).

Признаки, характерные только для человека
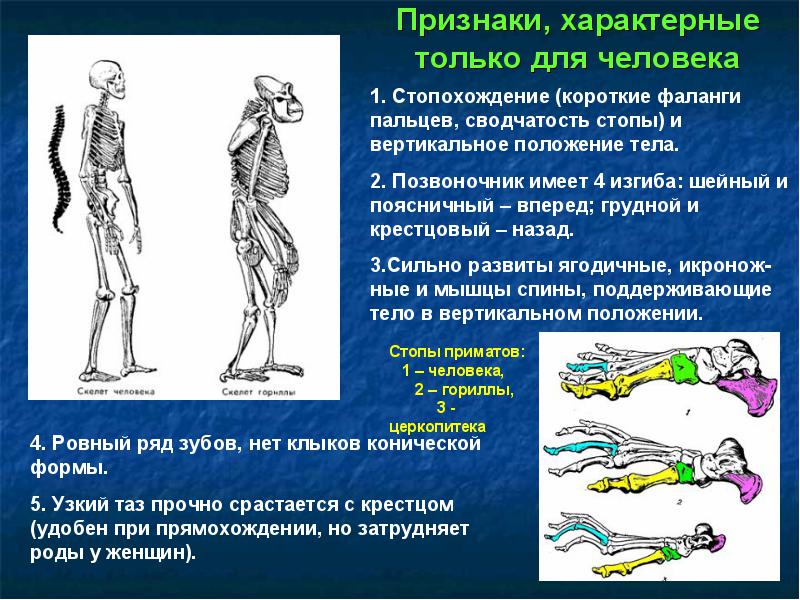
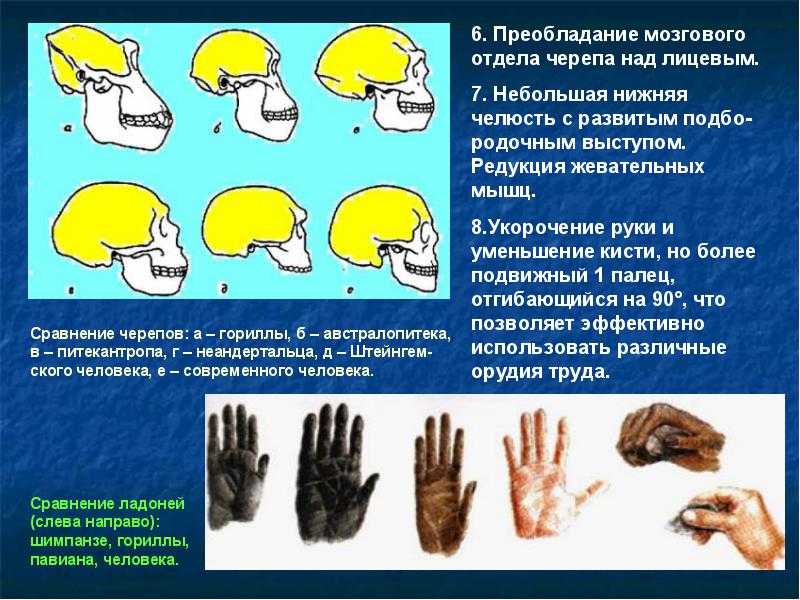
9. Развитый головной мозг, особенно лобная, теменная и височная доли коры полушарий.
10. Перемещение затылочного отверстия черепа к центру его основания и редукция мышечного аппарата затылка.
11. Низкое положение гортани (повышает роль ротового резонатора, но прерывает дыхание во время еды).
12. Редукция волосяного покрова.
13. Асимметрия тела и функции полушарий.
14. Увеличение времени взросления и полового созревания.
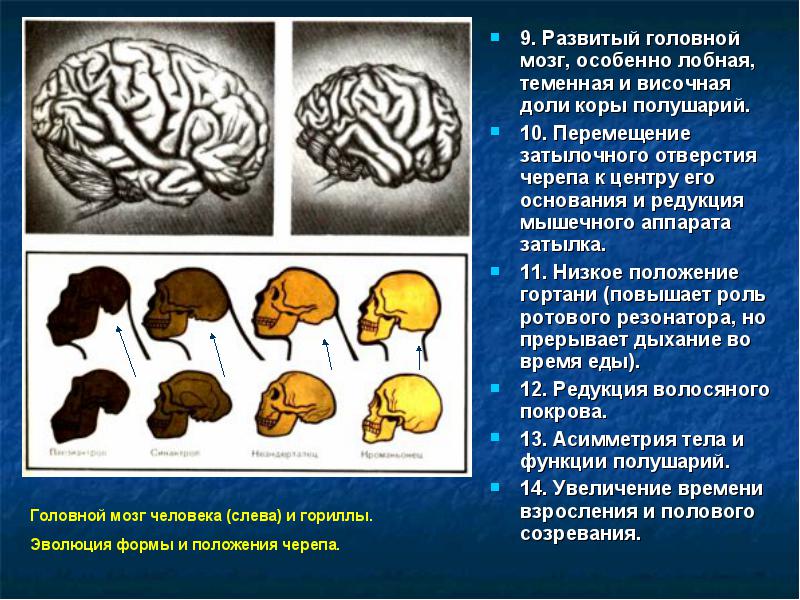
Изменение черепа в ходе эволюции гоминид
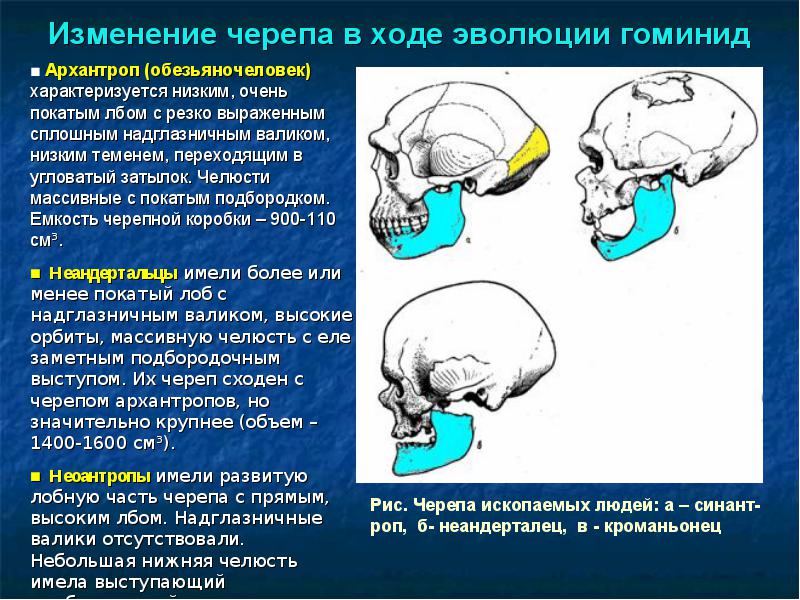
Отличие человека от животных
Разум (абстрактное мышление, а у животных мышление всегда конкретное).
Речь (животные общаются с помощью системы сигналов, но только у человека есть вторая сигнальная система – общение с помощью слов. Речь возникла в процессе труда).


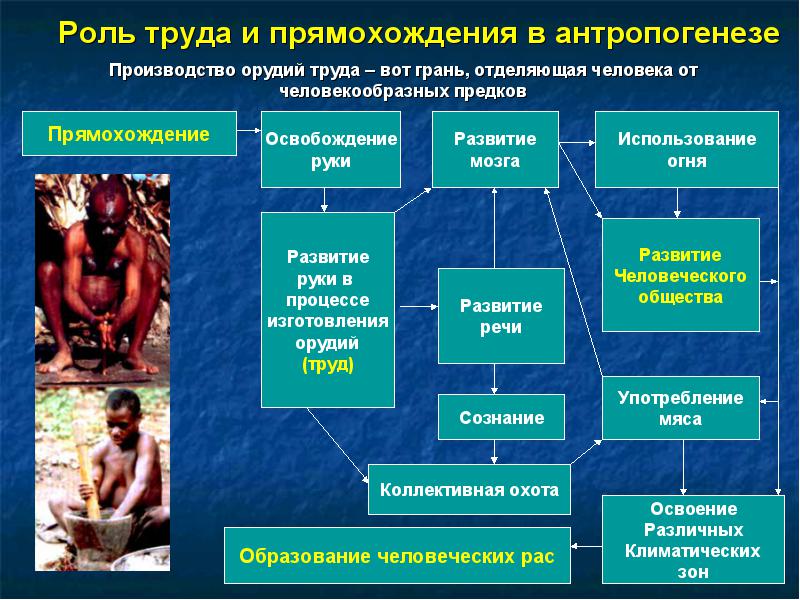

Расы
Раса – исторически сложившиеся в определенных географических условиях группы людей, обладающих некоторыми общими наследственно обусловленными морфологическими и физиологическими признаками.

в виде небольшой общины
Согласно новейшим научным данным, современное человечество имеет единых генетических предков в виде небольшой общины , обитавшей где-то в Центральной Африке. Генетические исследования

Классификация рас
Согласно классификации Я.Я Рогинского выделяют три большие расы: австрало-негроид-ную, или экваториаль-ную; европеоидную и азиатско-американскую, или монголоидную.
В пределах больших рас различают более мелкие группировки – подрасы, также отличающиеся друг от друга по ряду признаков.
Между большими расами и ареалами их распространения имеются промежуточные расы, сочетающие черты обеих рас, но в сглаженной форме.
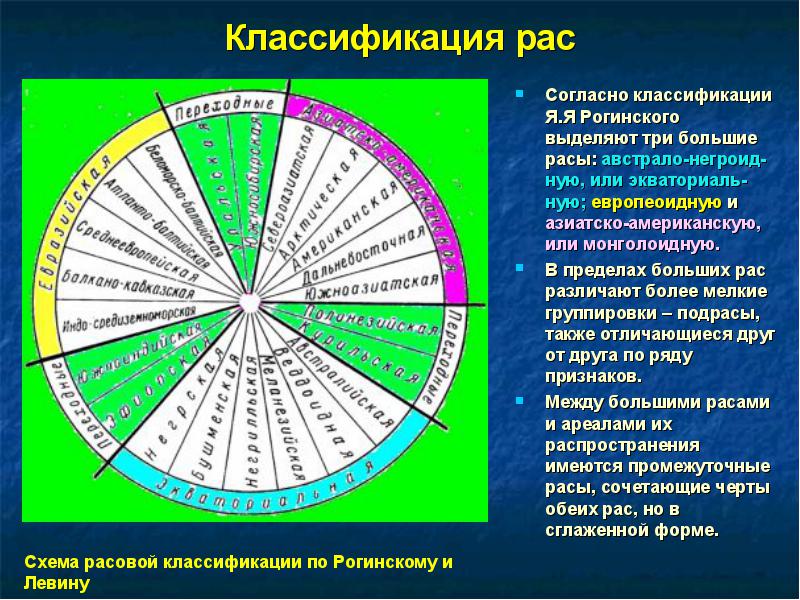
Европеоидная, или евразийская раса
Светлая кожа (разные оттенки).
Волосы мягкие, волнистые или прямые, от светлых до брюнетов.
Обильный рост бороды и усов у мужчин.
Лицо узкое, резко выступающий узкий нос, губы узкие.
Глаза горизонтально расположены, пигментация - от светло-голубых до черных.
Уши средней длины, со свободной, сросшейся или полусвободной мочкой
Ареал распространения: Европа, Передняя и Средняя Азия, Северная Африка. 40% населения Земли.
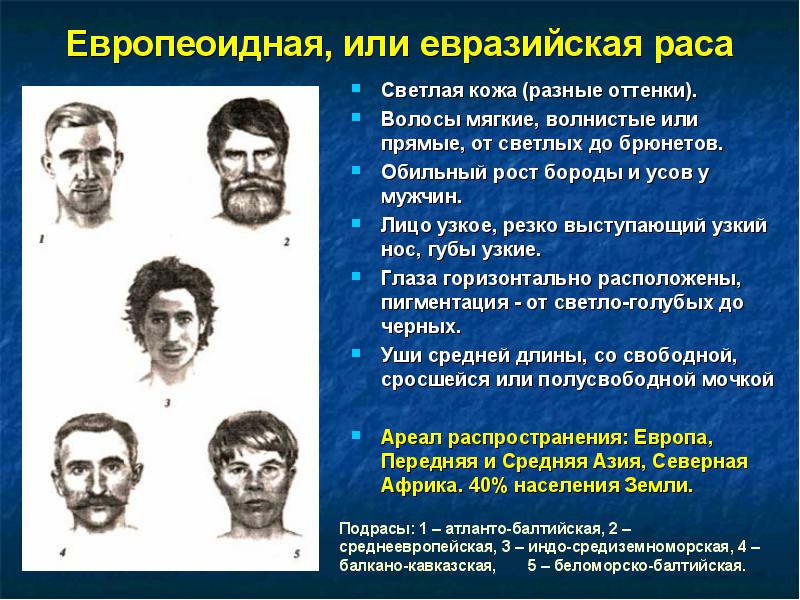
Азиатско-американская раса
Желтовато-смуглая, различных оттенков кожа.
Жесткие прямые черные волосы. Слабый или очень слабый рост бороды и усов у мужчин.
Лицо широкое уплощенное, скулы сильно выступают; нос уплощен (у американской подрасы – сильно выступающий), губы средней толщины.
Глаза раскосые, узкие, с характерной складкой на верхнем веке – эпикантусом; пигментация – темная.
Уши длинные со свободной мочкой.
Ареал распространения: Северная, Центральная, Восточная и Юго-Восточная Азия, Американские индейцы. 50% населения Земли.

Экваториальная раса
Темная кожа (от серо-сизой до коричнево-черной).
Черные волнистые или курчавые волосы, средний рост (обильный рост у австралийской подрасы) бороды и усов у мужчин.
Лицо чаще узкое с выступающей челюстной частью, широкий, с низким переносьем, мало выступающий нос, губы толстые.
Глаза широко открытые, темные.
Уши короткие со сросшейся или полусвободной мочкой.
Ареал распространения: Африка южнее Сахары, Австралия, Новая Гвинея, Зондские, Филиппинские острова и Меланезия. 10% населения Земли.

Переходные расы
Пограничными между тремя большими расами считаются:
1 – Эфиопская раса (между экваториальной и евразийской).
2 – Южноиндийская раса (между веддоидной и индо-средиземноморской).
3 – Уральская раса (между беломорско-балтийской и североазистской).
4 – Южносибирская раса (туранская) (между евразийской и монголоидной).
5 – Полинезийская раса (между монголоидной и экваториальной).
6 – Курильская раса (между австралийской и арктической).

Типы телосложения у разных рас
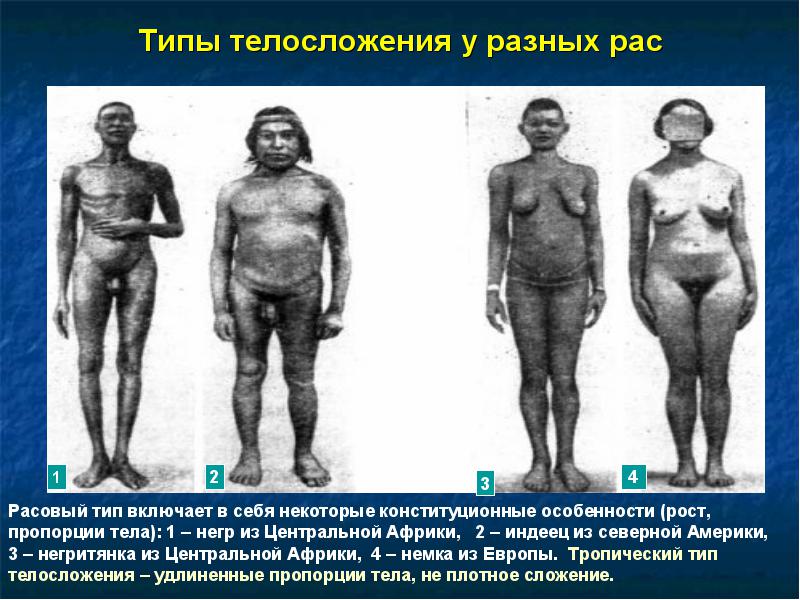
Расовые признаки черепа
Череп Евразийской расы узкий, с вертикально высокой лобной костью, выступающими носовыми костями, глубокой «собачьей ямкой» (клыковидной) на верхней челюсти.
Череп Монголоидной расы характеризуется слабым выступанием лица с широкими скуловыми отростками. Клыковые ямки не выражены. Носовые кости выступают слабо. Лобная кость не высокая, покатая.
Череп Экваториальной расы имеет широкие, слабо выступающие носовые кости, сильно выступающие вперед челюсти (прогнатность).

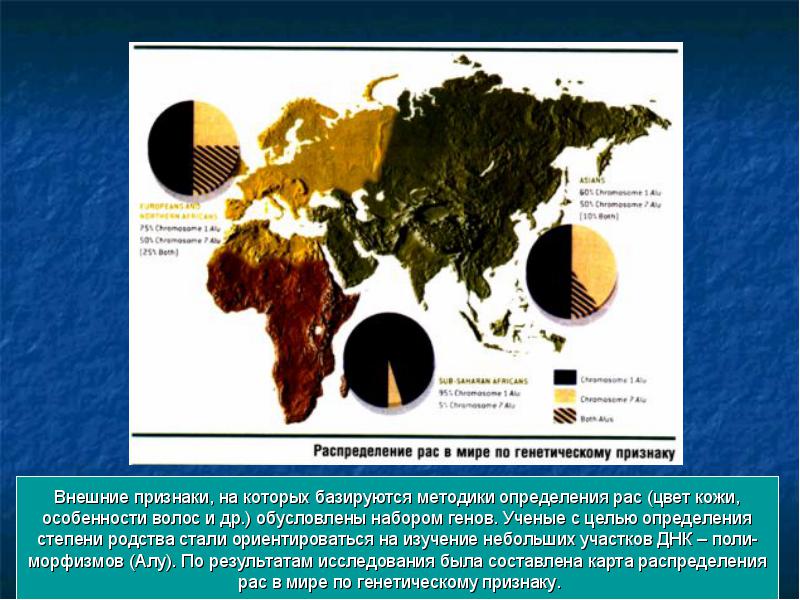
Происхождение рас
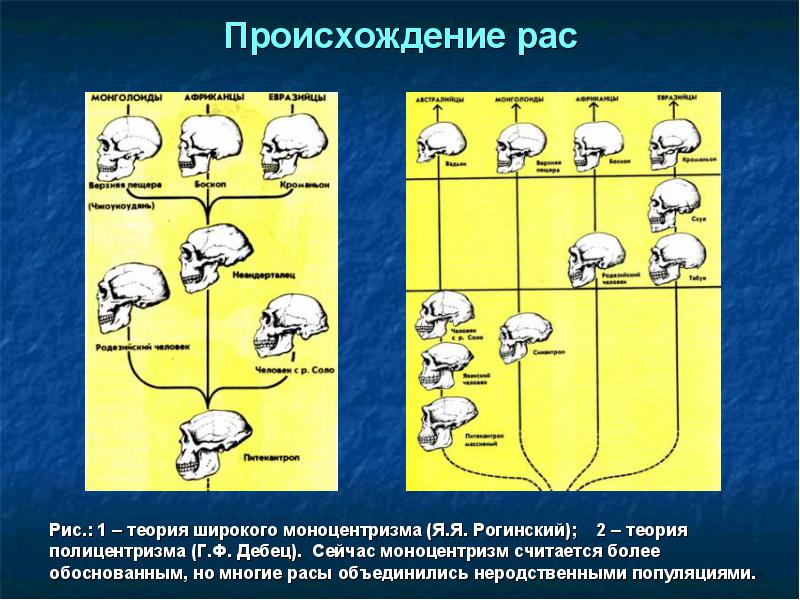
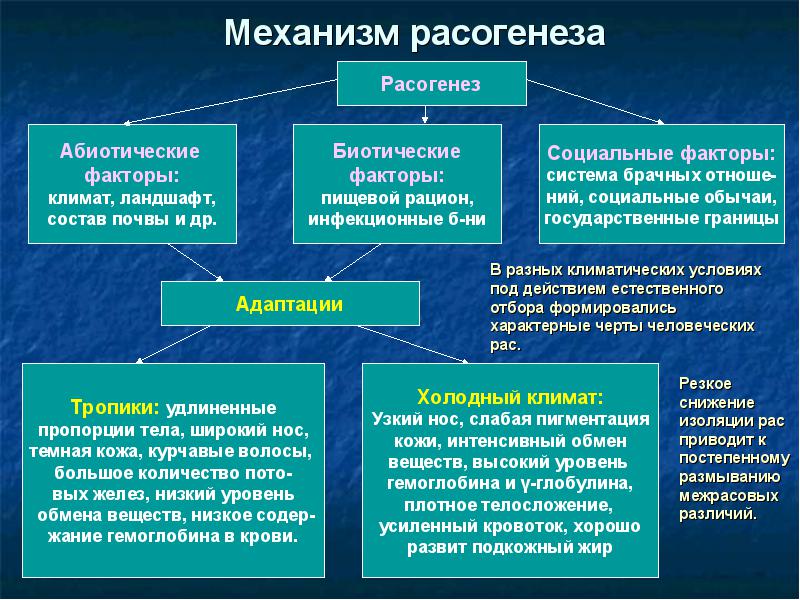
Современные первобытные охотники


морло,
В пустынной части Австралии обитают аборигены морло, называющие себя древними или истинными людьми. Они не имеют постоянного места жительства, т.к. ежедневно кочуют в поисках пищи. Они очень тесно связаны с Природой, поэтому никогда не остаются без еды и умеют находить воду там, где нет ни малейших признаков влаги.

Контрольно - обобщающий тест
1. Атавистические признаки у человека – это: а)густой волосяной покров , б)хвост, в)копчиковая кость, г)короткие мягкие волосы на теле.
2. Главный признак, впервые отделивший в ходе эволюции человека от других приматов: а)прямохождение, б)трудовая деятельность, в)речь.
3. Кто из перечисленных ниже приматов наиболее близкий родственник человека? А)мартышка, б)горилла, в)шимпанзе, г)орангутанг, д)гиббон.
4. Какой признак свидетельствует о биологическом единстве всех человеческих рас? А)сходная норма реакции на условия среды, б)одинаковый набор хромосом, в)плодовитость потомства при смешанных браках.
5. Назовите последовательность развития человека: а)неоантропы, б)архантропы, в)палеоантропы, г)современные люди, д)австралопитеки.
6. К древнейшим людям относят: а)питекантропа, б)человека умелого, в)синантропа, г)неандертальца, д)гейдельбергского человека, е)человека с о. Флорес.
7. Объем мозга кроманьонца составлял: а)около 1400 см3, б)около 1600 см3, в)800-1200.

9.Представители H.sapiens – это: а)питекантроп, б)синантроп, в)неандерталец, г)кроманьонец.
10. Какие из признаков человека не наследуются? А)речь, б)дыхание, в)мышление, г)коллективный труд, д)цветное зрение.
11. Чем человек отличается от человекообразных обезьян? А)наличием 4 групп крови, б)наличием речи, в)наличием грудных молочных желез, г)наличием сводчатой стопы.
12.Среди перечисленных факторов эволюции человека назовите биологические: а)трудовая деятельность, б)речь, в)развитие мозга, г)наследственная изменчивость, д)освоение огня, е)половой диморфизм, ж)прямохождение.
13.Зачаточная членораздельная речь появилась у: а)питекантропа, б)кроманьонца, в)неандертальца, г)человека умелого.
14. Общественный образ жизни у предков человека способствовал: а)появлению прямохождения, б)освобождению руки, в)появлению речи.
15. Останки палеоантропа впервые были обнаружены: а)в пещере Кро-Маньон, Франция, б)на о. Ява, в)в Олдувайском ущелье, Танзания, г)в пещере Неандерталь, Германия.
16. Культура созданная прегоминидами называется: а)неолит, б)мустьерская, в)ашерская, г)мезолит, д)олдувайская.
17. Укажите ископаемых предков человека, обитавших на Земле 9-15 млн. лет назад: а)питекантропы, б)проконсул, в)дриопитек, г)австралопитек, д)неандерталец.

18. Человеком прямоходящим называют: а)австралопитека, б)питекантропа, в)неандертальца, г)кроманьонца.
19. Какой признак, в отличие от человекообразных обезьян, присущ только человеку? А)трудовая деятельность, б)забота о потомстве, в)наличие 4 групп крови.
20. Какая коренная раса живет на севере Африки? А)экваториальная, б)евразийская, в)монголоидная
21. Культура, созданная архантропами, называется: а)ашерская, б)мустьерская, в)неолит, г)олдувайская.
22.Какие данные свидетельствуют о высоком развитии неоантропов и об окончании биологической эволюции человека?
23. В чем состоит ложность теории расизма?
24.Как вы объясните возникновение человеческих рас?
25. Укажите основные адаптивные признаки экваториальной расы?

Эталон ответов на контрольный тест
1. а,б; 2. а, 3. в, 4. в, 5. д,б,в,а,г; 6. а,б,в,д; 7. а, 8. в,г; 9. а,г; 10. б,г; 11. в,г,е,ж; 12. в, 13. в, 14. г, 15. д, 16. в, 17. б, 18. а, 19. б, 20. а.
21.Строительство жилищ, создание совершенных орудий труда из разнообразных материалов , развитая членораздельная речь, одомашнивание животных и выведение культурных растений. Все это привело к независимости человека от внешней среды и приостановке биологической эволюции;
22. Человек – полиморфный вид, все расы которого имеют одинаковые основные признаки, отличающие человека от животных; все они способны к труду и творческой деятельности и способны усваивать огромный объем информации; при скрещивании дают плодовитое потомство с явлением гетерозиса;
24. Естественный отбор при развитии адаптивных признаков к различным местам проживания и дрейф генов; 25. темный цвет кожи, темные курчавые волосы, темные глаза, широкий нос и толстые губы, более массивная нижняя челюсть.
